 设为首页
加入收藏
设为首页
加入收藏
扩展资料
种群(population)
在一定空间范围内同时生活着的同种个体的集群,例如同一鱼塘内的鲤鱼或同一树林内的杨树。 population一词源于拉丁语populus,原意为人群,后在生物学中推广至一切物种,乃译为种群,另外还有居群、繁群等译名。当用 population一词专指种群的数量时,则视具体物种的不同而有人口、兽口、虫口等名称。
种群一词与物种概念密切相关。根据生物学种定义同一物种的个体不仅因其同源共祖而表现出性状上的相似(包括形态、大分子结构及行为等各个方面),而且它们之间能相互交配并将其性状遗传给后代个体。但不同种之间则由于形态、生理或行为上的差异而不能交配繁育,这称为生殖隔离。与此相应,广义的种群即是指一切可能交配并繁育的同种个体的集群(该物种的全部个体)。例如世界上总人口。但在生物学上更关心的却是实际上进行交配繁育的局部集群,下面主要讨论的便是这种狭义的种群。
因此,同一物种可有许多种群分别存在不同地区。主要是地理的原因阻止了它们之间的交配。这地理的原因有两种,一种情况是存在着地理屏障,如岛屿上的兽群被海隔绝,绿洲中的兽群被沙漠包绕。这一类型的地理隔离是明显可见的。另有一种情况,如欧、亚、北美北部的广阔林带可绵延千里,其间环境条件连续渐变,无法找出明显界线可借以区分出个别种群。但林带中相距较远的同种个体仅因距离关系无法进行交配,这是另一类型的地理隔离。
上面这种建立在生物种概念上的种群定义主要来自对昆虫、鱼、鸟、兽等营有性生殖的动物的研究,它不适用于专营无性生殖的生物,如细菌。事实上,就是应用到植物,也出现一些问题。一般动物成体的大小、体重、以及器官和附肢的数目或相近或相同,但在同种植物个体间则变异甚大。由代谢角度来看,同种植物个体的生物量相差悬殊。每个植株吸收的阳光和养分以及它们合成的有机物质的量的差别可以倍计。而某些行营养繁殖的植物如竹、芦苇,可蔓延扩展覆盖广大地面。这些植体相连应视为单一个体,但其新生部分扎根地下,将它断裂却可独立成活。类似情况也存在于某些营固着生活的低等动物如许多腔肠动物。显然,将这样差异显著的个体都视作类似的种群的单位是不恰当的。因此另有人提出组件生物的概念,认为这些营固着生活的动植物是由称为组件的重复单位(如腔肠动物的螅型体或高等植物的枝条)组成的。因而在分析这些组件生物的种群组织时需要同时考虑个体及组件两个水平。
在生物组织层次结构中,种群代表由个体水平进入群体水平的第一个层次。因为有性生殖过程是一个基因重组过程,重组产生新的变异,可供自然选择,所以相互交配繁育的种群便构成了一个进化的单位,它可能成为分化新物种的起点。有的生物还环绕着繁育关系组成一定的社群结构。另一方面,同一地区的个体共享同一资源,因而在对待资源的关系上又表现出种内竞争或合作的关系。
种群内部的遗传过程是种群遗传学(现通常译为群体遗传学)的研究主题,种群成员间以及它们与环境之间的相互作用则为种群生态学的中心内容。这两者共同构成种群生物学。在理论上,种群研究与进化机制的探讨密切相关;在实用上,人口控制、生物资源利用、以及有害生物防治等问题实质上都是种群问题。因此,种群研究日益受到各方面的重视。
种群特征及其变化
种群特征指同种生物结成群体之后才出现的特征,因此大部分是数量特征,正因如此,在种群研究中常需要借助统计学。
种群的大小及密度
对界限明显的种群可以统计整体。但对一般种群则常常测定其密度,即单位面积或体积中的个体数量。在种群生态学中研究种群的物质代谢以及种群与自然环境的交互作用时,主要便是从密度着眼。但在群体遗传学中研究各种遗传性状在种群中的分布及其变迁时,却必须考虑互相交配繁育的整个种群。
种群大小的测定
除极少数物种的种群小且个体大易于观察外,对大多数物种都不必完全计数。因此常采用抽样方法加以估计。对于静止的生物如植物、土壤生物昆虫和农作物病虫害等,可选取一些具有代表性的区域──样方,计数一定面积或体积中的个体,再根据样方在总面积或体积中的比例推算出总体数字。为了选取确具代表性的样方,还需要先初步了解该物种在空间中的分布类型:是均匀型、聚集型、还是随机型。对于活动的生物,根据需要还可采用另一类抽样方法,例如标记重捕法。对第一次捕捉到的动物(第一次样本)标记后释放,经过一段时间估计标记动物已在总体中均匀分布后,再次捕捉并计数其中标记动物所占比例,根据第一样本数目(原标记数)及第二次样本中的标记比例即可推出总体数字。上述这些方法得出的都是绝对数字,但有时绝对数字难以得到,则可以调查数量的相对变动(或增加或减少)。这时也可以只测计一些间接的指标,如听动物的鸣叫声、观察地面粪便量的变化据此可粗略估计某种动物的数量增减。
种群的组成
同种生物虽具有相似性,但其具体遗传性状则个个不同。同种生物共有的大量基因决定了它们的相似性。此外还有很多基因在个体间是不同的,这些基因和每个生物面对的不同环境因子共同决定了它们之间的差异。在种群内,这些不同的基因中每一个只出现于一定比例的个体中,这个比例即为基因的出现频率。不同基因的组合(基因型)更是多种多样,每一种基因型也只存在于一定比例的个体中,这个比例即为该基因型的出现频率。这两种频率值反映了种群的遗传组成,都是群体遗传学的研究内容。在种群生态学中研究得较多的是种群的年龄组成和性别组成。它们是决定种群兴衰(数量变动)的重要因素。性别组成指雌性和雄性在种群中所占的比例,它是由性染色体的分布所决定的,因此也是一种遗传组成。一般说来,雌性生育子代,所以种群中雌性数量增多时,种群数量趋向上升。极端的例子见于营孤雌生殖的生物如蚜虫,它们在全年大部分时间中只有雌性存在,因而种群增长速度极快。年龄组成的意义在于,生物只在一定的年龄范围内才具有繁殖能力,而到达一定年龄则趋向死亡。因此,繁育年龄的个体在种群中的比例左右种群的出生率,老龄个体在种群中的比例影响种群的死亡率,而种群 出生率与死亡率的对比则决定种群的兴衰。
种群的结构种群并非同种生物的简单聚合。在繁育关系上,在资源利用上,它们常形成一定的结构,这在较进化的动物类群中表现得最为明显。例如在鸟兽中可以见到不同的交配体制:有乱交的、一夫多妻的、一夫一妻的等多种形式。一般说来,稳定的配偶制有利于保护幼体,有利于子代通过向双亲学习而增加适应环境的能力。社群结构常常是环绕着繁育关系建立起来的,但是一定的社群同时也是一个互相协作共同利用资源的单位。资源常按一定的优势顺序在社群内分配。很多动物在进入繁育时期常占据一定的领域,这种划分资源的行为常有助于保护自然资源使不致因抢食而枯竭。
种群的变动
种群的大小总在不断变动着,这决定于两组数字的消长:种群可以因出生或迁入新个体而增长,也可因死亡或迁出旧个体而减少。因生死造成的变化常称为自然增减;因迁入迁出造成的变化常称为机械增减。对于相对隔离的种群,其增减主要取决于出生率与死亡率的对比。出生率、死亡率、迁入率、和迁出率是研究种群动态的主要参数。在理想环境中(理化条件适宜、资源丰富、空间无限,无天敌相扰),一个物种能达到的出生率称为最大出生率或生理出生率,其数值较为固定。在实际环境中达到的出生率则称为实际出生率或生态出生率,其数值因具体条件而异。由于环境的限制,任何生物也不能在自然条件下实现其最大出生率,而昆虫等小动物的种群常随天气的波动而发生大幅度的增减。但一些大动物的种群大小却可在较长的时期内保持相对稳定,这说明存在着自动调节的机制。从理论上讲,当种群密度增加时,死亡率也相应增加或出生率相应降低,或两种情况同时出现,这都可以使种群密度维持稳定。这种因密度而变的因子称为密度制约因子。已发现,死亡率常是密度制约的,例如食源有限,个体多时只有强者得食而存活,弱者则受饥而死,因而死亡率随密度增加。这时出生率的下 降也可能是密度制约的,因当营养缺乏时,生育力也常随之下降。
种群间的相互作用和种群的进化
进化意味着种群中有利性状的增加和不利性状的减少,这自然是相应基因频率变化的后果。如果由于进化某个种群与同种的其他种群间出现生殖隔离,这就意味着新种的形成。由此可以看出,进化代表种群的质变,而对种群进行遗传学和生态学的综合研究正是探索进化机制的主要途径。基因频率和基因型频率的变化一方面是因为基因突变和重组提供了新的生物变异,另一方面则是自然选择的结果。在各种选择因子中,以生物因子最为活跃。捕食者必需捕获猎物才能生存,而猎物只有避开捕食者才能存活。在进化过程中,捕食者不断提高捕食能力,猎物则力求改善其避敌技巧。在寄生物―寄宿主系统中也有类似情况。总之,每一个种群是另一种群的选择因子,相互作用大大地促进了进化的速度,这种在相互作用下双方共同进化的情况称为协同进化。种群间相互作用也是种群生物学的重要内容。
(蔡晓明)
扩展资料
种群数量变动的因素
在自然界,决定种群数量变动的基本因素是出生率和死亡率,以及迁入和迁出等。出生和迁入使种群数量增加,死亡和迁出使种群数量减少。如果增量大于减量,种群数量则增加,相反时则减少,如果增量与减量相等,则维持不变。
1.出生率与死亡率
种群出生率(natality)是种群在单位时间内所出生的后代个体的百分数。例如,在一个有1000个个体的种群中,一年内出生了200个后代,这个种群的年出生率就是20%。理论上最大的出生率等于种群的繁殖力或繁殖潜能,即在理想的最适条件下种群不受外界因素限制时的出生率。但事实上,永不可能有理想的最适条件。并且由于生存竞争等的存在,繁殖力总要受到多方面的抑制。例如,由于竞争而出现的食物缺少以及气候的变动等都能影响生殖力(如影响卵的发育)。因此不是所有的个体都能产最多的卵,也不是所有的卵都能孵化或长成成体。所以实际的出生率或称生态出生率(ecological natality)总是低于理想的最大出生率的。
出生率的大小与性成熟的速度、胚胎发育所需的时间,每胎的卵或幼仔数以及每年繁殖的次数等有关。各种生物的生殖力有很大的差异。一般说来,低等动物的生殖力都高于高等动物。
死亡率(mortality)是种群在单位时间内死亡个体占种群总个体数的百分数。理论上最小的死亡率是指只有年老而自然死亡时的死亡率。但实际死亡率或生态死亡率(ecological mortality)总是远远大于最小死亡率。因为种群中大多数个体不可能都生活到它的生理寿命,总是要因疾病、饥饿、冻死、被捕食以及各种意外事件而夭折,这些都造成死亡率的增加。一般说来,种群密度增加时,死亡率一般也增大。当然,个体的寿命是决定死亡率的一个重要因素。各种生物的寿命也有很大的不同。昆虫多数是几天到几十天死亡,有的树木可以活几千年。
死亡对种群来说不一定是坏事,这必须与出生率联系起来一同看待。因为一些个体死亡了,在种群中留下空隙让出生的一些新个体来取代。这样的种群往往生活力更强。具有高死亡率、寿命短、但具有强生殖力的种群往往比一个长寿命的种群有更大的适应能力。
物理因素能影响出生率和死亡率。例如,一次寒冬的低温可以引起大量越冬昆虫的死亡,而一个适合的气候条件则可招致害虫的大发生等。
2.其他作用因素
除出生率和死亡率外,种群结构本身的特点如性比、年龄分布等,也都能影响种群数量。种群中如果雌性个体多的种群出生率显然要比雄性个体多的种群高。年龄分布也一样,生长快的种群,年轻个体多;衰退的种群中,老年个体多;稳定的种群具有比较均匀的年龄分布。所以可以由每一种群内年龄组的相对分布来说明该种群的数量的增长趋向。
动物的行为如扩散、聚集与迁移等可间接影响种群数量。例如,扩散使种群密度下降,因而使对种群密度起作用的控制因素(如疾病的传染、食物竞争、生殖力降低等)不起作用,种群得以继续增长数量。相反,聚集使密度增加,因而对密度起作用的控制因素起了作用,抑制了进一步增长,甚至于增加死亡率。迁出、迁入与扩散、聚集的作用是一样的。
所以种群数量对于多数生物来说,虽然可以达到稳定,但从来不可能是完全稳定不变的,因为自然界中永远存在着上述的各种物理的和生物的因素。
陈阅增,普通生物学,高教出版社
扩展资料
种群数量的调节
世界上的生物种群大多已达到平衡的稳定期。这种平衡是动态的平衡。一方面,许多物理的和生物的因素都能影响种群的出生率和死亡率,另一方面,种群有自我调节的能力,通过调节而使种群保持平衡。
1.密度制约因素和非密度制约因素
影响种群个体数量的因素很多。有些因素的作用是随种群密度而变化的,这种因素称为密度制约因素。例如,传染病在密度大的种群中更容易传播,因而对种群数量的影响就大,反之,在密度小的种群中影响就小。又如,在密度大的种群中竞争强度比较大,对种群数量的影响也较大,反之就较小。有些因素虽对种群数量起限制作用,但作用强度和种群密度无关。气候因素就是这样,刮风、下雨、降雪、气温都会对种群的数量产生影响,但这种因素起多大作用与种群密度也是无关的,这类因素称为非密度制约因素,无论是密度制约因素还是非密度制约因素。它们都是通过影响种群出生率、死亡率或迁移率而起着控制种群数量的作用。
2.密度制约因素的反馈调节
生物种群的相对稳定和有规则的波动和密度制约因素的作用有关。当种群数量的增长超过环境的负载能力时,密度制约因素对种群的作用增强,使死亡率增加,而把种群数量压到满载量以下。当种群数量在负载能力以下时,密度制约因素作用减弱,而使种群数量增长。现举几例说明这种反馈调节。
(1)食物 旅鼠(Lemmas)过多时,它们在草原大面积地吃草,草原植被遭到破坏,结果食物缺乏(加上其他因素,如生殖力降低,容易暴露给天敌等)。种群数量从而减少,但数量减少后,植被又逐渐恢复,旅鼠数量也随着恢复过来(图19-12)。
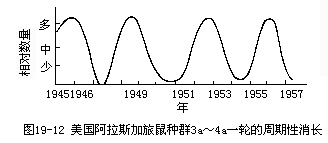
(2)生殖力 生殖力也受密度的影响,池塘内的椎实螺在低密度时产卵多,高密度时产卵就少。大山雀在英伦三岛的林区,每窝产卵数随密度的大小而减少或增多。这个效果也可能是由于密度高时食物缺少或某些其他因素的作用所引起的。
(3)抑制物的分泌 多种生物有分泌抑制物来调节种群密度的能力。蝌蚪密度高时产生一种毒素,能限制蝌蚪的生长,或增加蝌蚪死亡率。在植物中,桉树有自毒现象,密度高时能自行减少其数量。细菌也有类似的情况:繁殖过多时它们的代谢物就将限制数量的再增加;密度降低时,这些代谢产物少,就不足以起抑制作用,因而数量又能上升。
(4)疾病、寄生物等是限制高密度种群的重要因素 种群密度越高,流行性传染病、寄生虫病越容易蔓延,结果个体死亡多,种群密度降低。种群密度低了,疾病反而不容易传染了,结果种群密度逐渐恢复。
3.非密度制约因素的作用
生物种群数量的不规则的变动往往同非密度制约因素有关。非密度制约因素对种群数量的作用一般总是很猛烈的,灾难性的。例如,我国历史上屡有记载的蝗灾是由东亚飞蝗(Locusta migra-toria manilensis)引起的。引起蝗虫大发生的一个物理因素是干旱。东亚飞蝗在禾本科植物的荒草地中产卵,如果雨水多,虫卵或因水淹或因霉菌感染而大量死亡,因而不能成灾,只有气候干旱蝗虫才能大发生,所以我国历史上连年干旱常同时伴随虫灾。
物理因素等非密度制约因素虽然没有反馈作用,但它们的作用可以为密度制约因素所调节,即可以通过密度制约因素的反馈机制来调节的。当某些物理因素发生巨大变化(如大旱、大寒)或因人的活动(如使用杀虫剂)而使种群死亡率增加,种群数量大幅度下降时,密度制约因素如食物因素就不再起控制作用,因而出生率就得以上升,而种群数量很快就可恢复到原来的水平。
研究生物种群数量变动的规律和影响数量变动的因素,特别是种群数量的自我调节能力,就有可能制定控制种群数量的措施,对种群数量变动进行预测预报,为生产服务(如制定防治害虫的规划,对害虫、害兽发生的测报,以及决定狩猎与采伐的合理度等)。
陈阅增,普通生物学,高教出版社
扩展资料
群落组织问题
关于群落的性质问题,历来有两种对立的观点。一种认为群落是真实的实体,它是组成群落的各个物种种群相互作用形成的有组织实体。另一种认为,群落在自然界中并非是一个实体,而只是生态学家从一个呈连续地变化着的植被中搜集来的一组生物而已。两种观点争论的实质在于群落到底是一个有组织的系统,还是一种纯粹偶然的个体集合,有的学者形象地描述为“有序”(order)还是“混乱”(chaos)之争。前一种的极端代表认为,群落是一种超有机体,后一种的极端认为,群落纯粹是一个人为的、不切实际的抽象。因此,持前一种观点的可称为机体论学派(organismic school),而后一种则称为个体论学派(individualistic school)。
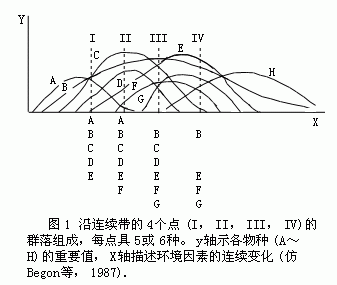
长期来,多数群落生态学家持机体论观点,如Clements、Tansley和Elton,所以绝大多数教科书把生物群落定义为一定生境中各生物种群相互作用而有机结合的结构单元。但持个体论学者从未中断争论,尤其是50年代以来,一些采用定量方法(如梯度分析和排序等)的研究证明,陆地群落并不是一个个分离的、有明显边界的实体,而是在空间和时间上连续的一个系列(图1)。即各物种在环境梯度上的分布彼此独立,而不是许多物种经常地结合在一起形成一个有机的物种群。大量排序研究结果都与此类似。这一事实使更多的生态学家倾向了个体论学派的观点,并占了上风。但机体论学派并不由此而停止争论,仍以各种变型和论点出现。还有一些学者认为没有一派的观点能包括全部真理,并提出已经到了停止争论的时刻了。
生态学研究有个体、种群、群落和生态系统等层次。个体,即有机体,边界很明确,其各个部分有机地组成整体也很明显,高等的还有神经内分泌系统进行协调。种群各部分相互作用虽不如此明显,但多有雌雄成幼、家族,甚至等级和社会组织,更重要的是通过基因的世代传递使种群保持为一整体。群落则不然,一般说来,边界比较模糊(往往具过渡性的群落交错区),内部结构也比较松散,至今未找到组织群落的明确的内部机制,这正是导致机体论和个体论两派长久之争的原因。
1989年美国国家科研委员会出版的《生物学中的机会》一书中认为:“群落生态学中最令人感兴趣的问题是为什么有这么多的动植物种数,为什么它们现在这样分布着,以及它们是怎样发生相互作用的。……到什么程度群落才确实是整体结构,而不是简单的个别种的聚合……”。许多当代群落生态学家认为,群落生态学的中心问题就是群落的整体结构是如何形成的,其机理如何,即所谓的群落组织(communityorganization)。群落组织这个概念与群落结构有所不同,后者强调形态,多是描述性的,前者强调形成整体结构的机理,深入到规律性问题。
就目前群落学发展的水平而言,学者至多仅能说明群落的一部分组织,主要是通过种群间相互作用有机结合成的“物种包”(speciespacking),或称群落的亚集。下面介绍其一些进展。
(1)竞争对群落结构的影响。
大概没有人怀疑竞争在形成群落结构中的作用,同样也不会有人认为群落中所有物种都是由种间竞争而联结起来的,所以问题的焦点在于:在形成群落结构的因素中,竞争起多大作用,在什么条件下其作用大,什么条件下作用小。
种间竞争通常多出现在生态位比较接近的种类之间,如食果鸟类、食虫蜥蜴类等,对此1967年Root提出同资源种团(guild,或译功能团、共位群)概念,例如食果鸠鸽就是一同资源种团。Brown(1975)曾分析北美荒漠中食种子啮齿类,在一个荒漠中有5种,平均体重分别为7.1、12.5、18.1、37.6和101g。Hutchison曾据此类结果提出:共存物种间通常以“标准量”而分开。他这里所指的“标准量”是共存物种间个体大小的比率,即较大物种大约是较小物种的1.5~2倍。这就是说,食性相同的物种由于激烈的竞争,造成生态位分化(在此例中个体大小分化,各食不同大小种子),这些同资源种团中的物种,通过竞争而得以共存。
对种间竞争在形成群落结构中的作用,最直接证据是通过引种或去除(某种)试验。例如在Arizona荒漠中有一种更格卢鼠和3种囊鼠共存,食性和栖息小生境上彼此有些区别,当去除一种,另3种利用的小生境都有扩大。
Schoener曾总结文献中164例这类试验,平均有90%例子证明有种间竞争。植物是自养生物,都一样要求光、水、CO2和营养物,研究种间竞争和生态位分化有较大困难。但Tilman模型及其实验研究,同样说明竞争在形成群落结构中的重要作用。
(2)捕食对群落结构的影响。
如果捕食者食性很广,例如野兔,实验证明,随着兔食草压力的加强,草地上植物种数有所增加。其原因是兔把竞争力强的物种个体吃掉,可以使竞争力弱的物种更好生存,所以多样性提高。但是,当食草压力过高,兔不得不食适口性低的植物,植物种数又随之降低。因此,植物多样性与兔捕食强度的关系呈驼背状曲线,它表明中度捕食下,物种多样性最高,此即中度捕食假说的根据。潮间带滨螺捕食藻类,试验研究同样证明中度捕食假说。
海星是岩底海岸群落的捕食者,它以贻贝、帽贝、藤壶等为食。Paine通过人工去除海星,证明去除后物种数目由15种降为8种。由此证明顶极捕食者是维持该群落多样性的关键,学者称它为关键种(keystone species),关键种决定了群落的面貌。
(3)干扰与群落结构和动态。
干扰是平静的中断,或正常过程的妨碍,干扰是自然界的普遍现象。连续的群落由于干扰而出现中断(可称为断层,gap)是常见现象。如森林中由于大风、雷电、大象、伐木、疾病,可出现倒木,形成林窗。Cornell等根据断层出现频率对物种多样性的研究,提出中度干扰假说,即中等强度的干扰能维持高多样性。其理由是:①一次干扰后少数先锋种入侵断层,如果干扰频繁,一再出现,则先锋种不能发展到断层的演替中期,使多样性保持较低;②如果干扰间隔期很长、干扰频率低,演替能发展到顶极期,竞争排斥作用达到排斥别种,多样性也不很高;③只有中等干扰程度将使多样性最高,允许更多物种入侵和建立种群。这情况与捕食强度与物种多样性相似,实际上,捕食也是一种干扰。
断层的另一种后果是出现抽彩式竞争。珊瑚礁中鱼类特别丰富,对如此高的多样性,人们难以仅用食物资源分隔解释得通,实际上,许多种鱼食性是相同的。礁中有许多小空隙,据观察,在原有领主死亡后空隙为哪种鱼占据完全是随机的,没有出现有规律的演替。由此可见,礁中高多样性的维持,也决定于许多相同生态位的物种对空间的抽彩式竞争。当群落中由于各种原因不断出现新的断层,时而在此,时而在彼,时而为这一种中彩,时而为那一种中彩,群落整体的多样性就比没有干扰和断层情况下更高。
(4)空间异质性与多样性。
群落的环境一般不是均匀一致的,而具有空间的异质性,例如小地形、土壤水分等都不均匀。空间异质性的程度越高,意味着有更多样的小生境、更多样的小气候条件,更多样的躲避天敌的隐蔽所,所以能允许更多的物种共存,包括竞争共存(通过资源分隔)和非竞争共存(通过更广的资源范围和更复杂的环境条件)。例如,Harman研究了淡水软体动物与空间异质性的相关,他以水体底质的类型数作为空间异质性的指标,得到了正的相关关系:底质类型越多,淡水软体动物种数越多。植物群落研究大量资料,也说明土壤和地形变化丰富的群落含有更多的植物种数。
通过以上介绍使我们看到,早期的群落生态学是描述性的,描述群落的结构和演替过程,强调群落的分类;近代的群落生态学焦点在研究形成群落结构的机制,其研究方法也强调了实验研究和模型研究或理论生态学研究,正如Schoener所指出的,群落生态学的机理性研究途径。群落生态学中最令人感兴趣的问题是群落中为什么有这么多种数的动植物,为什么它们像现在这样分布着,以及它们是怎样发生相互作用的。机体论者强调群落的整体性,个体论者强调群落性质决定于各个物种,这是多年来的争论。这种非此即彼的争论也许使答案模糊而延误了进展,更有说服力的观点也许是把现实的自然群落视为个体论――机体论轴(或称Gealson-Clements轴)中的任何一点,即由种间相互作用紧密性不同程度变化的连续谱中的一点。实际上,群落的原生演替过程就是由一个简单到复杂、由种间联系不紧密到紧密的发展过程。近代的群落生态学文献中一种群落分类的新标准,把群落分为无相互作用和有相互作用群落(non-interactive和interactive communities)两类(它们是连续谱的两端),正反映了这种发展趋势。
孙儒泳,生物学通报,1993年第5期
扩展资料
引进生物新品种的利与弊
由于地理,地貌,气候等原因,世界上动植物种类的分布有明显的区域性,此地盛产之物,彼地就成为罕见之物。因此,古今中外的科学家们,为各国各地区引进生物新品种做了大量的工作,但这里有成功的喜悦,有失败的教训;有得到收益的兴奋,也有引起祸害的辛酸……。
我国从西汉张骞出使西域时开始,陆续从国外引进苜蓿、葡萄、蚕豆、核桃、大蒜、石榴、马铃薯、玉米、辣椒、花生、红薯、南瓜、棉花、西瓜、菠菜、茄子、姜、芝麻、豌豆、西红柿、荞麦等农作物,以及沉香、菩提树、贝叶树、悬铃木、油橄榄、白杨、黑松、冷杉、橡胶树、桉树、银合欢、相思树、南洋杉等树种。同时,我国的水稻、大豆、茶、油桐、桃、杏、柑桔,以及多种名贵花卉,特产珍树也在异国的土壤中开花结果。我国的四不象(麋鹿)还是首先由我国传入英国,后在我国绝迹,到本世纪50年代再由欧洲返回家园繁衍后代的。由此可见,生物引进的意义重大,它不但丰富了各国人民的生活,而且使世界上的动植物家族更加繁荣兴旺起来。
然而,在几千年的引进史上也不乏失败的例子,更有甚者,由于盲目引进而引起无穷的祸害。
在生物引进史上造成危害最大的恐怕要数烟草的引种了。1492年,哥伦布发现美洲新大陆,当他们看到当地人口中叼着一卷燃着的干草棒时,初时惊叹不己,继而羡慕之极,后耒竞相效尤。1558年,烟草在哥伦布的家乡西班牙试种,随后迅速蔓延欧洲各地,并逐步走向世界。明朝时,烟草经菲律宾传入我国。烟草对人类的危害可谓罄竹难书,要是当初人们将它拒之门外,也就不会有无数的瘾君子死于它的魔掌之下了。
对我国恐怕最令人辛酸的是英帝国主义强迫我国引种鸦片(罂粟植物)。1840年以来,英国殖民主义者强迫我国人民种植鸦片,结果有许多人成为吸毒者,害得家破人亡,同时中国的几亿两白银和大量财富随之流入英国侵略者的腰包。
在作物的引种上,古今中外也出现过不少失败的事例。汉武帝刘彻曾因荔枝“味道好极了”于公元111年便下令在长安建造一座“扶荔宫”,从当时的“蛮夷之地”引种了100株荔枝,结果竟全部冻死,虽一再补种,仍无一幸存。尽管武帝“怒发冲冠”杀了几十个无辜的养护人,也无法使荔枝在长安生存,最后只能“偃旗息鼓”。解放后的50年代,我国曾在东北引种南方良种水稻,结果苗株茂盛但不抽穗扬花,而南方引种东北水稻也同样颗粒无收。
不顾植物本身对特定环境的适应性,盲目引进失败固然可笑,而不顾本地实际情况盲目引种“成功”却十分可怕。有一种生长在南美洲的仙人掌,在当地虽然生长快,但因为有的动物以它为食,所以没有造成任何危害。但自从被引种到澳大利亚,由于它飞快繁殖,把大片大片的牧草地全部“占领”。人们慌了,用铲子铲,用锄头锄,用拖拉机深翻,都奈何不了它,后来多亏生物学家想了一种办法,从它的老家带回一种大蝴蝶,它的幼虫专吃仙人掌,才使仙人掌的生长得到控制,保住了牧场。无独有偶,一种生长在巴西的鳄鱼草,被北美和非洲一些国家引种后,肆无忌惮地狂长,长满了海湾,湖泊,池塘;长满了各个角落,怎么也消灭不了。后来只好到巴西搬来“救兵――一种专吃鳄鱼草的跳甲虫,才控制了它的灾害。
大米草既可护堤,喂牛,又可用作燃料。1983年福建省把它当作宝贝从美国引进,在东吾洋海滩推广种植。过去面积14万亩的东吾洋,不仅是多种鱼类的天然鱼库,而且是全国对虾、贝类的养殖基地,滩涂生物达200多种。但是自从大米草在此“安家落户”后,不到几年它们盘根错节,生长茂盛,布满整个水区。海水涨潮时滩涂生物被冲进草丛无法逃生,以致许多水产品,如蛏、蛤、章鱼、跳鱼等,濒临绝迹;海水退潮时,浮游生物附着滞留在草丛中,致使人工养殖的海带,紫菜,牡蛎、对虾等,因海水缺乏营养而产量锐减,造成了巨大损失。时至今日,仍无法清除。
在生物引进史中,动物引进的数量相对植物而言可谓“九牛一毛”。然而,引进不当,危害比植物毫不逊色。最著名的恐怕是澳大利亚的“兔害”和“癞蛤蟆事件”了。
1859年,澳大利亚一位庄园主托马斯・奥斯金在英国游玩时,看到本国没有的小动物――兔子十分可爱,于是带回24只放养在自己的庄园里。想不到,兔子在澳大利亚迅速繁殖生长,在不长的时间里便遍布全国各地。它们破坏植被,引起土壤侵蚀,啃光大片牧草和庄稼。据统计,因兔子造成的经济损失每年高达1亿美元以上。生物学家曾多次设法对付兔害,但均未成功。本世纪50年代,曾培育出一种能使兔子不育的跳蚤,一度使兔子死亡90%以上;然而到80年代初,这里的兔子已对跳蚤病毒产生了免疫力,于是兔子家族又迅速兴旺发达。目前,澳大利亚至少有兔子4亿只以上,灾害日益严重。
刘德明,生物学教学,1994年第6期