 设为首页
加入收藏
设为首页
加入收藏
扩展资料
病毒从何而来?
病毒从何而来?这是生物进化问题上一个至关重要的研究课题,是一个不折不扣的悬而未决的问题。特创论者们常常在一些情况不甚明朗的生物学领域中兴奋无比,在病毒起源上也如同其它方面那样喋喋不休地歌颂着那个被古闪米特人造出来的“伟大的上帝”。而科学家们则在苦苦地探索着病毒的起源及其演化过程,以求将它们定位在大分子的起源与不同的病毒宿主出现之间的进化树上的某一位置。但这是个极其困难的工作,因为根本就没有病毒化石用来做证据,这迫使科学家们去另辟途径,最有代表性的做法是从病毒与细胞的相互关系方面寻找蛛丝马迹。
核酸含磷元素而不含硫元素,蛋白质含硫元素而不含磷元素,根据这一特点,可进行这样一个实验:
1.让培养基含有32P(磷放射性同位素之一),植入大肠杆菌;
2.细菌开始生长,并被32P做了标记;
3.加入噬菌体(细菌病毒),使其感染被32P做了标记的大肠杆菌;
结果:在A培养基中得到许多被32P标记的噬菌体。
4.将被32P标记的噬菌体放入未被作标记的大肠杆菌并使后者感染;
5.进行搅拌后,做低速离心;
结果:在离心沉淀(大肠杆菌细胞)中测到大部分放射性,而离心上清液中则只有很少量的放射性。
6.改用35S(硫放射性同位素之一)重做上述1─5步骤搅拌实验,
结果:在离心沉淀(大肠杆菌细胞)中测到很少量放射性,而离心上清液中则有大部分的放射性。
实验表明:病毒感染进入细胞的只是它的核酸部分,而病毒的蛋白质外壳留在了细胞外。
以上这个经典的“搅拌试验”是由Herhey和Chase两位科学家完成的。
一般说来,病毒入侵细胞并进行活动有这要样几种情况,一是杀死细胞来换取病毒自身的增殖,最著名的例子是嗜菌体;二是感染细胞后不杀死细胞而增殖;三是感染后不增殖即处于一种隐匿状态,后面两种情况最具代表性的是多瘤病毒,这类病毒对小鼠细胞感染后不杀死细胞而增殖,对大鼠细胞则为隐匿状态。从这里我们可以得知不同的病毒在不同细胞中的行为有很大的差异,换句话说,病毒与细胞的关系远比我们想像的要复杂得多,而这种多元化关系又给我们研究病毒带来了极大的困难。进入20世纪90年代后,随着对病毒与细胞相互作用的分子模型分析和病毒核酸的分子生物学研究以及病毒基因克隆技术的发展,人们对病毒的起源形成了三种代表性的假说。
第一种假说:病毒是地球上生物进化过程中最为原始的生命物质,它产生于化学进化之后,因此它既有化学大分子属性,又具有生物的部分特征。该学说暗示病毒是从无生命到有生命的过渡型物质,其位置处于化学大分子和原始细胞之间。地球生命演化的过程表现为:无机物→有机物→化学大分子→病毒→原核生物→真核生物。这个假说是根据生命起源学说和分子进化理论所提出来的,一种纯粹的假设,缺乏任何进化上的证据。
第二种假说:病毒是高级微生物的退行性生命物质,即微生物在其生存过程中可能会丢失部分基因,这部分基因丧失了独立的自我繁殖能力,只能在重新进入微生物细胞中找到相应的位置才能具有活力,久而久之便退化为病毒。这种生命形式让我们立即想到了立克茨体和衣原体,它们比细菌小且更原始,它们只是专性细胞内寄生的中间形式,因此本假说认为这是病毒形成的依据,其过程为:细菌→类似立克茨体类的生物→类似衣原体类的生物→病毒。对于这个假说最不利的情况是我们无法找到在细胞内寄生的小型细胞生物,并且在立克茨体和衣原体中未发现病毒。故这个假说成立的证据不足。
第三种假说:病毒来源于正常细胞的核酸,因偶然途径从细胞内脱离出来进而演变为病毒。该假说又称内源性假说。其间接的实验证据为:
1.作为细胞一部分的质粒可随时脱离细胞,并在细胞间传递,病毒与质粒是相似的。
2.相当多的DNA病毒的DNA能全部或部分结合到它们所寄生的细胞的染色体上,从而成为细胞的一部分,这正好是细胞核酸外逸的逆过程。
3.正常细胞中存在较广泛的逆转录型可动遗传因子和逆转录型重复序列,提示正常的细胞中含有RNA所介导的DNA合成反应,而这与逆转录病毒的核酸的复制行为相一致。
这个假说的间接证据可部分解释 DNA 病毒的起源,但要说明RNA病毒的起源则相当困难。
由于1999年在古细菌中发现了所谓“反转子”的遗传单位,这为病毒在基因水平上的起源及进化提供了新的证据:“反转子”是仅含一个基因并且能自我复制的一段核酸分子,具有重要的基因捕获功能。其基因宿主细胞等位基因高度同源,且在密码子的使用频率上也有高度的相似性,经过考证,初步估计“反转子”在4亿年前就存在了。在古细菌进化的过程中,反转子从细菌基因中捕获基因,扩大自身的遗传信息量并增加生物学功能,最原始的感染性病毒颗粒由产生,原始病毒采取不同的复制策略,因此产生了不同病毒的进化。由此生命的起源是从细菌开始的,细菌病毒进化摇篮。据此推测:地球上最早出现的病毒可能是经RNA为基因组的逆转录病毒,然后再发展成以DNA为基因组的逆转录病毒,最后出现的是DNA病毒,而RNA病毒则很可能有其独立的演化途径。
总而言之,病毒起源是复杂和多元化的,不同病毒有着不同的起源,至少在DNA病毒和RNA病毒之间是如此,随着更多研究资料的积累,相信我们会获得更多的新证据。有一点目前是有基本的共识的,那就是病毒是一个可动遗传单元,在相应的条件下进行细胞之间的定位传递。作为大分子物质,病毒起源实际就是生物大分子的起源,是当今生命科学中最重要的命题之一,乘着本世纪生命科学的全面发展之翼,这方面应该有重大的突破。
来源:三思科学
扩展资料
32亿年前的细菌
发现在非洲南部的单独曙细菌化石是迄今为止科学家发现的最古老的细菌化石,也是所有古生物化石中最古老的代表。单独曙细菌是一种原核生物,年代测定表明的生活时代为距今32亿年前。由于类似于单独曙细菌这样的地球上的最早的生物类型都是结构很原始的单细胞生物,即使形成化石也非常轻散、脆弱、易碎,因此长期以来,科学家一直没有发现这些原始生命的其它可靠的化石。
后来,一些科学家在对水成岩中的风化型条带状富铁矿的成因进行分析时,竟然发现这种富铁矿是由一种生活在远古的微生物--铁细菌形成的;而且,形成这些富铁矿的那些铁细菌生存的年代最早也可以上溯到32亿年前。
铁细菌具有一般细菌的共同特征,都是直径只有几微米到几十微米的单细胞生物,而且是细胞内没有成形细胞核的原核生物,只有在放大成千倍的显微镜下才能发现它们。有些铁细菌细胞为椭圆形或杆形,相互联系起来形成相当长的线体,有的单个铁细菌就是一条细而长的线体;有些铁细菌呈球形、弧线形或杆形带柄或分枝的形态;有的铁细菌形成小瘤状、带状或螺旋状。这些铁细菌外面都包裹着一层薄薄的“铁甲”――皮鞘。
铁细菌在生活过程中,摄取铁质和硅酸等无机物。在沼泽和湖泊中,铁元素通常以可溶性的氢氧化亚铁的形式存在,被铁细菌慑入后,在菌体内经过酶的催化作用,把它氧化成不溶性的三氧化二铁。
这些不溶性的铁化物和硅化物等无机物被铁细菌分泌到体外,就形成了以铁为主要成分的皮鞘。十分有趣的是,铁细菌的皮鞘往往比其身体大几倍或几十倍。铁细菌可以在皮鞘中前后移动,有时还可以伸出鞘外,重建新的皮鞘,而脱落的皮鞘就在水中沉淀下来,聚集成铁矿。你可能不会想象到,这种生活在亿万前年的铁细菌,竟是通过这样的生活方式,成了造铁的“能工巧匠”,为今天的人类提供了极为丰富的铁矿资源。
在美国、加拿大、苏联、澳大利亚、印度和非洲南部前寒武纪距今18亿年到32亿年前的沉积岩中,科学家都发现了条带状的铁矿层,其中普遍含有铁细菌化石。如果将岩石或矿石磨成薄片,在高倍率的生物显微镜或电子显微镜下观察,就可以看到铁细菌化石。
由于对远古时代的铁细菌化石的研究还不够,现在仅发现十多种。它们和现代的铁细菌有些相似,但又有所不同。其中有的种已经绝灭。当然,那时候的种绝不等于现代的种,例如,线状古铁细菌与现代线状铁细菌很相像,它们都有皮鞘,但是古代的比现代的大得多。
铁细菌大多是好氧微生物,但又需氧不多。科学家推测,32-34亿年前的地球大气具有高度的还原性。那时,大气中几乎没有氧气存在,仅有二氧化碳、甲烷和氢气。到了32-31亿年前,蓝细菌出现了。在其后直到27亿年前的岁月里,这些蓝细菌摄取原始的还原性大气中的二氧化碳,通过叶绿素进行光合作用,产生出游离的氧气并释放到大气中,使大气中的氧气含量逐渐增加,这就为好氧的铁细菌提供了良好的生活条件,从而保障了它们能够不断地摄取水中的氢氧化亚铁并把之氧化成不溶于水的三氧化二铁沉积到水底。
可见,今天常常引起人们恐慌和误解的细菌,它们遥远的祖先不仅奠定了整个生物界在以后的岁月里进化发展的生物学基础,而且其中的一些分子还为发展到今天的智慧生物--人类的进步积累了不可或缺的矿物资源。
扩展资料
引起种群基因库组成变化的原因
突变、选择、迁移以及偶然因素能引起种群基因频率变化,它们是小进化的主要因素。
如果不存在上述因素,则一个有性种群的遗传组成保持相对恒定。换句话说,在种群内不发生突变,种群成员没有迁出,也没有其他种群成员迁入,没有自然选择作用,没有任何其他进化因素作用的情况下,有性生殖过程不会改变种群基因库的基因频率。这就是遗传学中的哈代-温伯格平衡(Hardy-Wenberg equilibrium)原理。
以一个位点上的等位基因为例:假设一个最简单的情况,即考查种群某一位点上的两个等位基因A和a,它们的频率分别为p和q。种群内三种基因型个体的频率分布见表7-1。
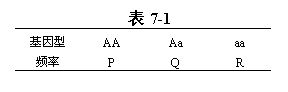
假定种群内个体之间随机互交,则各交配组合的频率分布见表7-2。
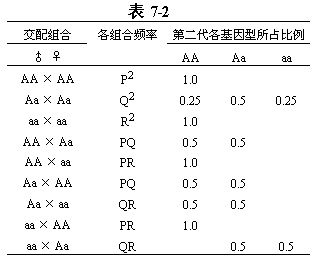
第二代基因型AA的频率:
P′=P2+0.25Q2+0.5PQ+0.5PQ
P′=P2+PQ+0.25Q2
P′=(P+0.5Q)2
由于P+0.5Q恰为等位基因A的频率p,所以第二代基因型AA的频率为(P+0.5Q)2=p2。同样的程序可知,第二代基因型aa的频率为q2,基因型Aa的频率为2pq(参考Ridley,1993)。
于是可知,有性生殖种群在无突变、无迁移、无选择的情况下,一对等位基因A和a经过一代有性生殖后的频率变化见表7-3。

由上表可知,假定第一代亲本基因型为AA和aa,等位基因A和a的频率分别为p和q(p+q=1),有性生殖产生的第二代个体有AA、 Aa和aa三种基因型,其频率分别为p2、2pq和q2,第二代的等位基因A和a的频率实际上没有变,仍然是p和q。这表明,有性生殖虽然使个体不能将其基因型不变地延续到下一代,但整个种群基因库的基因组成并不因有性生殖而发生变化。了解这一点对了解小进化是重要的。
引起种群遗传组成变化的第一个因素是基因突变。
如果种群内发生突变是随机的(准确地说,相对于种群的适应进化方向而言,突变的方向和性质是不定的),那么,突变本身并不造成驱动种群基因频率定向的改变,即不产生突变压;如果发生定向突变,例如等位基因A以每世代为u的突变率变为等位基因a,回复突变不发生,或回复突变率V<u,则突变本身构成驱动种群内基因频率定向改变的因素,即形成突变压。假设种群内等位基因A的初始频率p0=1,即全部个体的基因型为AA,经过n世代以后,等位基因A的频率由于突变而下降,这时A的频率
pn=p0(1-u)n
每一世代等位基因A的频率的改变量
Δp=p-p(1-u)=up
即等位基因A的频率以每世代-up的速率下降,此处的负数代表频率下降。另一方面,种群内等位基因a的频率q以每世代uP的速率增长。因此,突变可以看作是驱动进化的因素或进化的一个驱动力。突变率的高低可用来衡量这种力的大小,即通常所说的突变压。但高等动、植物的突变率低(每个基因每世代的突变几率为10-5数量级),单由突变压引起的种群基因频率的改变是很缓慢的。例如,假定突变率u=10-5,经过10代以后,等位基因A的频率pn=(1-0.00001)10,只减少0.5%。
引起种群遗传组成改变的第二个因素是种群成员的迁出和迁入。个体在种群间的迁移意味着种群间的基因交流,这会直接造成种群遗传组成的改变。改变的程度取决于迁移的规模和种群的大小。
单向的迁出,即种群成员的流失,也同时造成基因的流失。单向的迁入,即种群接受外来的个体,也同时引进新的基因。小种群发生较大规模的迁移时(无论是迁入还是迁出),种群遗传组成都会发生大的变动。
种群之间的双向迁移,即种群之间互有迁出和迁入,会引起种群间遗传差异的减少,种群内的变异量增大。
哈代-温伯格平衡原理只适用于大种群,而小种群中的有性过程往往造成遗传组成的飘移;环境波动、灾害以及其他偶然因素也可造成小种群遗传组成的大的变化。
造成种群进化改变的最重要的因素是选择(这里指的是自然选择)。选择是一个非随机因素,是种群遗传组成的适应性改变的主要原因。我们在后面要详细阐述。
引起种群遗传组成变化的其他因素还有繁殖方式,例如近交和远缘杂交。
在隔离状态下的小种群,自体受精的生物,运动能力有限和配子散布的限制以及婚配习性等都可能造成长期的近亲繁殖。长期近交的后果是种群遗传均一化,种群内变异量减少。这种状况有利于种群保持已获得的适应特征,却不利于适应变化的环境。
扩展资料
植物与动物的分异
按照生物的五界分类系统,植物和动物作为生物的两个高层次分类阶元,是分别从另一个阶元――原生生物界的一些不同门类中进化而来,而且与后者是呈并列关系的。
这样的分类系统突出了生物各大阶元之间存在的从简单到复杂、由低级至高级的层次关系。
但是,它也有不足之处。特别是没有反映现代生物的两个最基本和最进步类群――动物与植物的系统关系及其历史渊源。
实际上,植物与动物的祖先类型不仅都可以在原生生物中找到,而且它们在原始生物中的祖先类型甚至具有一定的同一性。这种同一性在现代的一种原始生物--眼虫身上还可以找到。
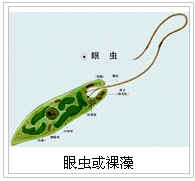 眼虫是一种生活在水中的单细胞原生生物。身体呈长梭形或圆柱形,前端有一个凹口,由此伸出一根鞭毛,其摆动在水中产生的反作用力能够推动身体运动;凹口的下方有一个具有感光机能的红色的眼点(眼虫的名称就因具有眼点而得)。如果把它们放在含有有机物的水中,眼虫能够靠细胞膜吸取水里的有机物“食物”,过着动物式的异养生活。这些性质使动物学家认为,眼虫是一种“原生动物”。但是同时,眼虫的细胞却又有含叶绿素的叶绿体,能够进行光合作用,自己制造营养。因此,植物学家认为,它是一种“原生植物”;由于它的细胞外面没有细胞壁,植物学家给它起了另外一个名字――裸藻。
眼虫是一种生活在水中的单细胞原生生物。身体呈长梭形或圆柱形,前端有一个凹口,由此伸出一根鞭毛,其摆动在水中产生的反作用力能够推动身体运动;凹口的下方有一个具有感光机能的红色的眼点(眼虫的名称就因具有眼点而得)。如果把它们放在含有有机物的水中,眼虫能够靠细胞膜吸取水里的有机物“食物”,过着动物式的异养生活。这些性质使动物学家认为,眼虫是一种“原生动物”。但是同时,眼虫的细胞却又有含叶绿素的叶绿体,能够进行光合作用,自己制造营养。因此,植物学家认为,它是一种“原生植物”;由于它的细胞外面没有细胞壁,植物学家给它起了另外一个名字――裸藻。
眼虫的这种“动物植物双重性”使许多科学家相信,动物与植物有共同的祖先――它很可能就是与眼虫类似的、某种生活在远古水域中的单细胞原生生物。在漫长的进化过程中,它们当中的某些个体伴随着基因组的变化加强了运动、摄食的结构和功能,同时逐步“丢失”了进行光合作用的结构和功能,最终生活方式转变成为完全的异养;另外一些个体则伴随着基因组的其它方式的变化向着完全自养的方式转变。前一种方式代表着最早的动物的产生,后一种方式代表着最早的植物的出现。
原始的原生动物和原生植物分异伊始都是单细胞的,随后,它们分别向多细胞方向发展。
在古生物学界,对植物与动物分异时间的认识是随着化石的不断发现与积累、新的研究思想、研究方法的进步以及学科交叉的相互影响而不断更新的。
早在达尔文于1859年发表《物种起源》之时,他就同时以实事求是的科学态度提出了当时的进化论所存在的难点,其中之一就是著名的“寒武纪爆发”。
所谓的寒武纪爆发是指科学家在距今5亿7千万年前到5亿年前的地层中发现的似乎是突然出现的众多的化石动物类群,其中包括海绵动物、腔肠动物、环节动物、软体动物、节肢动物、腕足动物、棘皮动物以及原始的脊索动物。根据这些发现,一些科学家认为这些动物在地球上是以一种爆发式的过程突然地出现在地球上的,同时,这种动物的爆发也说明了动植物分野的开始,因此他们推测,动植物分异的时间近于6亿年前的寒武纪之初。
到了1949年,古生物学家斯帕里格在澳大利亚南部阿得雷德山脉以北的埃迪卡拉地区发现了数量众多的无骨骼的海洋无脊椎动物化石,并把这一化石动物群的时代判定为寒武纪早期。可是10年后,古生物学家格拉斯南经过对这个动物群的认真细致研究,得出了三个不同凡响的结论:(1)这个化石动物群中无论是腔肠动物、环节动物还是节肢动物,它们当中都没有发现任何寒武纪的属种;(2)这个化石群中的微体化石组成与好望角的微体化石组成毫不相同;(3)埋藏着这个化石动物群的邦特岩层有1000米厚,与它上面覆盖的寒武纪地层之间并不连续(这在地层学上叫做不整合接触),因此邦特岩层应属于与寒武纪不同的地质年代。在1960年召开的国际地质学大会上,科学界正式把这个化石动物群命名为埃迪卡拉动物群。此后进行的多种手段绝对年龄的测定表明,埃迪卡拉动物群的年代为距今6亿8千万年至6亿2千万年。1974年,国际地质科学联合会将埃迪卡拉动物群确定为前寒武纪晚期的动物群。至此,古生物学家把无脊椎动物在地球上大规模出现的时间推前了,因此动植物分异的时间也被认为比6亿年前更加久远。
就在埃迪卡拉动物群被正确地重新认识的同时,以真核生物所形成的植物为主的植物化石的发现与研究也有了突破。1969年克劳德在美国加利福尼亚东部的贝克泉组地层中发现了生活于13亿年前的单细胞的绿菌和金藻化石;1971年舍夫和他的同事在澳大利亚苦泉组地层中发现了一些生存于9亿年前属于甲藻类、红藻类和绿藻类的植物化石。
70年代中期以来,前寒武纪的动植物化石的发现愈来愈多。一方面,已经发现的化石群在数量上和发现地点上都有增多,例如埃迪卡拉动物群现已经在西南非、北美、英国、斯堪的维纳半岛、苏联、中国等地的前寒武纪晚期地层中先后被发现,化石群的成员由最初的5个属发展到了19个属,到80年代初则已鉴定出了56个属。另一方面,新的化石群时有发现,例如中国学者在安徽省淮南地区的前寒武纪晚期地层中发现了丰富的须腕动物和环节动物化石,其标本和内部构造的清晰、完好程度在国内外都非常罕见,已鉴定出距今7亿4千万年前的须腕 动物化石和距今8亿4千万年的环节动物化石7个属。
须腕动物和环节动物都是属于高等的无脊椎动物,因此,学者们认为在此之前动物已有一段相当长的发展史,因此认为动植物的分野始于10亿多年前。
20世纪70年代以来,一些学者研究了蛋白质的分子结构。蛋白质是由氨基酸组成的化合物,许多氨基酸分子以链状的多肽连接在一起组成蛋白质。一个蛋白质分子由一个或多个多肽链组成。多肽链上的氨基酸排列顺序组成蛋白质分子的初级结构。蛋白质的初级结构--氨基酸的排列顺序,不但决定了蛋白质分子的二级、三级、甚至四级结构,而且从初级结构的差异可反映出不同物种之间的遗传差异和物种之间的亲缘关系。
迄今为止,科学家们已经弄清了数百种蛋白质分子的初级结构,所建立的分子进化系统与传统的分类系统基本吻合。这说明生物物种的分子进化和形态进化基本上是一致的。1982年,分子生物学家柳思尼考选择了在大多数无脊椎动物和全部脊椎动物中都存在的血红蛋白和肌红蛋白进行研究,结果表明无脊椎动物至少在10亿年前就已经出现了。另外一些学者对细胞色素C的研究则表明,地球上动植物的分异时间甚至远在13~12亿年前。
扩展资料
重返加拉帕戈斯
――正在进化的故事

达尔文雀
顽固的神创论者不但妄图否定进化论,而且还要否认进化这一事实,他们闭着眼睛说:“瞧啊,进化根本就不存在!”。那么,如何观察到进化呢?一个天文学家如果要了解恒星的演化,当然不可能等待上百亿年――没有那么长寿的天文学家。但是通过观察许多处于不同演化阶段的恒星,还是能够洞察天体演化的奥秘。生物学家可以通过化石记录来了解生物的进化。当然,更有效的方法是直接观察进化的过程――或许你认为这是一个非常漫长的过程,没有哪个进化论生物学家能够如此耐心和长寿。你错了,直接观察进化是完全可能的。普林斯顿大学的格兰特夫妇对于达尔文雀的研究就是一个很好的例子。
与鸟为伴
加拉帕戈斯群岛是太平洋赤道附近的一个群岛,在厄瓜多尔以西大约1000公里。几百年前,这个群岛还没有如此出名,甚至很少出现在航海图上。“加拉帕戈斯”在西班牙语中就是海龟的意思。现在,这个群岛简直是全世界生物学家心中的圣地,因为达尔文曾经在他的环球航行中考察过这个群岛。而且,这个群岛在很大程度上和他改变世界的科学理论联系在一起,那就是进化论。
1831年12月,达尔文乘坐英国海军贝格尔号军舰开始了为期5年的环球考察。1835年,他来到了加拉帕戈斯群岛。在这个群岛上,有着各种奇异的生物,例如巨大的海龟、蜥蜴和鸟类。达尔文注意到,加拉帕戈斯群岛上每一个岛屿的生物都与众不同,例如每一个岛上海龟壳的花纹和形状都有差别。达尔文还收集了加拉帕戈斯群岛上很多鸟类的标本――一个与世隔绝的群岛上居然有很多种外形不同的鸟类,它们的体形、喙以及生活习性的多样性让人觉得不可思议。
然而,在加拉帕戈斯群岛上看到这一切起初并未引起达尔文的高度重视。直到回到伦敦,他才逐渐发现加拉帕戈斯群岛上的这些奇形怪状的鸟儿其实是同一类鸟,它们同属于雀形目雀科。他甚至后悔没有更仔细的采集这些鸟类的标本。这种现象促使达尔文思考:它们很可能是源自同一祖先,在加拉帕戈斯群岛上漫长的生存过程中逐渐发生了变化。达尔文通过加拉帕戈斯群岛上鸟的变种认识到,物种并非永恒不变。这些小鸟在某种程度上促使达尔文产生了生物进化的思想。现在,人们习惯把加拉帕戈斯群岛上的这些小鸟称为达尔文雀。
达尔文雀不仅仅是放在博物馆中供人们参观和大发感慨的标本,它们也为研究生物的进化提供了绝好的机会。如果你要实际研究进化,那么最好找一个理想的实验场。这块试验场与世隔绝,不会有外界的生物进入,从而影响试验场内部生物的进化。加拉帕戈斯群岛正好符合这个条件。它们全都远离大陆,有人形容,加拉帕戈斯群岛的每一个岛都是动物们的自然监狱。动物们在岛上生老病死,一生都不会离开。
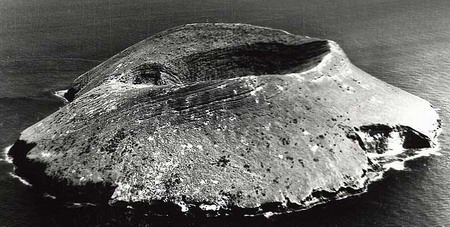
达芙尼岛:一个外形很像藤壶的火山岛
1972年,普林斯顿大学的生物学家格兰特夫妇来到这个著名的群岛,他们选择了其中的一个岛屿――达芙尼岛(Daphne Major)作为研究进化的实验场。这个岛荒无人烟,外形看上去就像达尔文曾经研究过的一种生物――藤壶。在那里,格兰特夫妇和他们的学生坚持观测了30年。他们记录下了岛上每一只达尔文雀,给它们带上脚环标记,记录它们的喙与体形,最多的时候他们能分辨出岛上2000多只达尔文雀。
在生死之间
格兰特夫妇观察到,即使是非常微小的因素,也能影响到达尔文雀的命运。达尔文雀最主要的区别体现在它们的喙和体形上。不同的喙适应不同的食物,例如大而钝的喙能够磕开最坚硬的植物种子,反之则适合小种子。当岛上的食物源发生变化的时候,鸟喙平均尺寸0.5毫米的偏差就能决定一种达尔文雀命运。自然选择促使了达尔文雀的外形不断变化。
达芙尼岛上主要有两种达尔文雀,一种是中喙地雀(Geospiza fortis),一种是仙人掌雀(Geospiza scandens)。中喙地雀的喙比较钝,它们以常绿植物的小种子为食。体形较大的中喙地雀还能吃非常坚硬的蒺藜种子。仙人掌雀的喙更尖,它们食用仙人掌的果实并为仙人掌授粉。仙人掌雀这个名字确实名副其实,因为它们把巢也筑在仙人掌上。

中喙地雀(Geospiza fortis)

仙人掌雀(Geospiza scandens)
在4月26日出版的《科学》杂志上,格兰特夫妇报告了他们对于达芙尼岛上两种达尔文雀的最新研究成果。他们发现,自然选择并非改变达尔文雀的唯一力量,两种达尔文雀之间的杂交也会使它们发生变化。
达芙尼岛的气候并非年年相同,例如1977年的席卷加拉帕戈斯群岛的拉尼娜现象造成了一场空前的干旱,岛上的小种子消失殆尽,大部分中喙地雀因为失去食物来源而饿死。当然,有一少部分喙较大的中喙地雀是幸运儿,它们的喙又大又钝,能够打开坚硬的蒺藜种子由于自然选择作用,中喙地雀的喙因此平均增大了4%。
世事难料,1983年的厄尔尼诺现象又让加拉帕戈斯群岛的降雨量充沛,各种植物变得非常茂盛,遍地都是小种子。命运的天平向反方向倾斜:喙比较大的中喙地雀吃这些小种子反而不如喙较小的中喙地雀有效率。自然选择又使中原地雀的喙平均缩小了2.5%。
自然选择也影响到了仙人掌雀。1983年的厄尔尼诺现象造成了空前的“洪涝灾害”。你知道,这对仙人掌可不是什么好消息。因此,仙人掌雀的食物来源也受到的影响。这次的幸运儿是那些喙稍钝的仙人掌雀。它们可使改吃植物的小种子渡过难关。我们也能料想到,这肯定促使仙人掌雀的喙向钝的方向变化。
但是格兰特夫妇吃惊的发现了一个悖论:在食物危机的自然选择压力消失以后,仙人掌雀的喙还是年复一年的变钝。这似乎是与常理相悖的。
最终,他们解决了这个“悖论”。1983年的大雨改变了仙人掌雀种群的性别比,雌性仙人掌雀大量死亡,雄性和雌性的比例甚至达到了五比一。无奈之下,部分雄性仙人掌雀和雌性中喙地雀交配,并产下了具有繁殖能力的后代。通常,两种达尔文雀之间的结合并不常见,即使这种情况发生,它们产下的后代也不具有繁殖能力。
中喙地雀和仙人掌雀的雄性杂交后代学习它们的仙人掌雀父亲求偶的歌声。因此它们也只选择仙人掌雀作为配偶。最终结果是,(雌性)中喙地雀的基因逐渐渗入了仙人掌雀的基因库,仙人掌雀的喙因此而变得越来越钝。这种现象称作“基因渗入”。
正在发生的进化
这一研究成果引起了生物学家的关注。因为这种现象很难在自然界的其他地方观察到,只有加拉帕戈斯群岛与世隔绝的环境下,经过长期而细致的研究才能揭示出其中的奥秘。
格兰特夫妇认为,这一研究成果表明,获得外来基因的物种可能更能适应环境的变化。同时,这种基因渗入现象也表明物种之间的隔离并没有人们认为的那么严密。
发生在加拉帕戈斯群岛上的进化故事正在、并且必将继续下去。格兰特夫妇的研究说明,短期的进化预报是可能的,但是长期预报就不太可能。气候的不确定性、两种生物之间基因的渗透都可能改变它们的命运。
在达尔文时代,加拉帕戈斯群岛上一共有13种达尔文雀。那么,若干年之后仙人掌雀会不会变得越来越像中喙地雀,从而消失在中喙地雀种群之中?彼得・格兰特说:“我认为这种融合现在就在进行中。”
来源:三思科