 设为首页
加入收藏
设为首页
加入收藏
扩展资料
氮的循环
氮是构成生物蛋白质、核酸的主要元素,因此它与碳、氢、氧一样在生物上具有重要的意义。氮的生物地球化学循环过程非常复杂,循环性能极为完善。氮的循环与碳的循环大体相似,但也有明显差别。虽然生物所生活的大气圈,其含氮量(79%)比含二氧化碳量(0.03~0.04%)要高得多,但是氮的气体形式(N2)只能被极少数的生物所利用。虽然所有的生物部要以代谢产物的形式排出碳和氮,但几乎从不以N2的形式排放含氮废物。在各种营养物质的循环中,氮的循环实际上是牵连生物最多和最复杂的,这不仅是因为含氮的化合物很多,而且在氮循环的很多环节上都有特定的微生物参加。
(1)固氮
由于大气成分的79%是氮气,所以氮最重要的储存库就是大气图,但是大多数生物又不能直接利用氮气,所以以无机氦形式(氨、亚硝酸盐和硝酸盐)和有机氮形式(尿素、蛋白质和核酸等)存在的氮库对生物最为重要。大气中的氮只有被固定为无机氮化合物(主要是硝酸盐和氨)以后,才能被生物所利用。虽然固氮的方法有物理化学法和生物法两种,但其中以生物固氮法最为重要。据估计,靠电化学和光化学固氮,每年平均可固氮7.6×106吨,而生物固氮平均每年的固氦量为54×106吨,人类每年合成氮肥约30×106吨,这也是一个不小的数字。根据人类合成氮肥的增产速度,预计到本世纪末,每年约可生产氮肥100×106吨。有些科学家认为:现在的工业固氮量约等于现代农业到来之前的生物固氮量。
固氮过程首先需要分子氮激活,使其分裂为两个自由氮原子,这个过程需要消耗能量,在生物固氮时,l摩尔的N2约耗能669×103焦。在自由氮与氢结合形成氨气时,l摩尔的氮气(28克)可释放54×103焦能量,因此,固定l摩尔的氮气,只需输入615×103焦能量就够了。除了光化学固氮法外,所有固氮生物部需要从外部提供碳化合物作为一种能源,以使影响这一吸热反应。生物固氨需要两种酶(固氮酶和氢化酶)进行调节,生物固氮的意义在于低能消耗,而工业固氮需要极高的温度和极大的压力(即400℃高温和200个大气压)。
已如有固氮能力的细菌和藻类很多,但为了方便可把它们分为两个类群:一类是共生的固氮生物(主要是细菌,但也有真菌和藻类),另一类是自由生活的固氮生物(包括细菌、藻类和其他一些微生物)。共生的固氮生物主要生活在陆地,而营自由生活的固氮生物在陆地和水域都有。但共生固氮生物在数量上至少要比营自由生活的固氮生物多几百倍。
在共生固氮生物中,根瘤菌是最重要的,也是人类了解最清楚的。根瘤菌对宿主植物(如豌豆、三叶草和菜豆等豆科植物)有高度的特异性,一定种类的根瘤菌只同一定种类的豆科植物发生共生关系,这些根瘤菌可潜入豆科植物的根毛,然后进行繁殖。已知有10多种高等植物(如鼠李、杨梅和桤木等)也有共生生物固氮作用。由于豆科植物与根瘤菌之间已经形成了密切的共生关系,所以豆科植物离开了根瘤菌就不能固氮,而把根瘤菌接种在其他植物是也不能固氮。
正如前面已说过的,在水生生态系统中,固氮生物大都是非共生生物,但有一个重要例外,这就是满江红及其共生生物蓝绿藻(鱼腥藻),它们广泛分布于我国温带和亚热带的水稻田中,被农民作为肥料加以利用,因此对农业生产有重要意义。在非共生固氮生物中既有需氧细菌也有厌氧细菌,还包括蓝绿藻。需氧固氮菌(如固氮菌属)广泛分布在土壤中以及淡水和海水中;厌氧菌(如梭菌属)的分布也同样广泛。事实证明:土壤和水体中的很多细菌都有固氮能力,又由于它们数量极多,所以它们固定的氮量也相当可观。例如在盐沼的沉积层中,细菌固氮量相当于藻类固氮量的10倍。这里值得强调的一点是:所有的共生和非共生固氮菌部需要从外部供应糖类,以使作为一种能源去完成固氮过程的吸热反应,因为没有任何一种固氮菌能够进行光合作用。
为了研究水体中的固氮过程,需把溶在水中的大气氮移出(通常是用氦清除),然后用一种稳定的氮同位素15N取而代之。并且质谱分析仪跟踪观察这种同位素的去向,这种方法与使用14C测定初级生产量的方法大体相同。研究表明:高固氮率与三种蓝绿藻(鱼腥藻属)存在着正相关。对其他两个湖的研究也表明:高固氮率与其他蓝绿藻的大量存在密切相关。而在马尾藻海,高固氮率则与束毛藻的存在相关。固氮过程所需要的能量是靠这些蓝绿藻的光合作用提供的,也就是说,蓝绿藻所生产的有机物质提供了固氮所需要的能量。
(2)氨化作用
当无机氮由蛋白质和核酸合成过程而形成有机化合物(主要是胺类)以后,这些含氮的有机化合物通过生物的新陈代谢又会使氮以代谢产物(尿素和尿酸)的形式重返氮的循环圈。土壤和水中的很多异养细菌、放线菌和真菌都能利用这种富含氮的有机化合物。这些简单的含氮有机化合物在上述生物的代谢活动中可转变为无机化合物(氨)并把它释放出来。这个过程就称为氨化作用或矿化作用。实际上,这些微生物是在排泄它们体内过剩的氮。有些具有氨化作用的微生物只能利用胨而不能利用简单的氨基酸,或者只能利用尿素而不能利用尿酸。相反,其他的微生物则能利用多种多样的含氮有机化合物。氨化过程是一个释放能量的过程,或者说是一种放热反应。例如,如果蛋白质的基本构成物是甘氨酸,那么l摩尔的这种蛋白质经过氨化就可释放出736×103焦的热能。这些能量将被细菌用来维持它们的生命过程。
(3)硝化作用
虽然有些自养细菌和海洋中的很多异养细菌可以利用氨或铵盐来合成它们自己的原生质,但一般说来,这些含氮化合物难以被直接利用,必须使它们在硝化作用中转化为硝酸盐。这个过程在酸性条件下分为两步,第一步是把氨或铵盐转化为亚硝酸盐;第二步是亚硝酸盐转变为硝酸盐。亚硝化细菌可使氨转化为亚硝酸盐,而其他细菌(如硝化细菌)则能把亚硝酸盐转化为硝酸盐。这些细菌全都是具有化能合成作用的自养细菌,它们能从这一氧化过程中获得自己所需要的能量。它们还能利用这些能量使二氧化物或重碳酸盐还原而获得自己所需要的碳,同时产生大量的亚硝酸盐或硝酸盐。研究表明,亚硝化细菌每同化一个单位的二氧化碳就可使35个单位的氨氧化为亚硝酸盐;而硝化菌每同化一个单位的二氧化碳可使76~135个单位的亚硝酸盐氧化为硝酸盐。硝酸盐和亚硝酸盐很容易通过淋溶作用从土壤中流失,特别是在酸性条件下。
目前,对开阔海洋及其海底沉积物中的硝化作用还不十分了解。1962年,首次报道了从开阔大洋海水中分离出来的海洋亚硝化菌,研究表明,这是一种专性自养细菌,它只能从氨中获得能量和从二氧化碳中获得碳。不少科学家认为,氮素是海洋浮游植物生产量的主要限制因素。
(4)反硝化作用(也称脱氮作用)
反硝化作用是指把硝酸盐等较复杂的含氮化合物转化为氮气、一氧化氮、一氧化二氮的过程,这个过程是由细菌和真菌参与的。这些细菌和真菌在有葡萄糖和磷酸盐存在时可把硝酸盐作为氧源加以利用。大多数有反硝化作用的微生物都只能把硝酸盐还原为亚硝酸盐,但是,另一些微生物却可以把亚硝酸盐还原为氨。在无氧条件下和有葡萄糖存在时,硝酸盐还原为一氧化氮的反硝化过程是一种放热反应,l摩尔的硝酸盐约可放热2.28×106焦。若将l摩尔的硝酸盐还原为分子氮,则可放热2.38×106焦。
由于反硝化作用是在无氧或缺氧条件下进行的,所以这一过程通常是在透气较差的土壤中进行的。依据同样的道理,在氧气含量很丰富的湖泊和海洋表层,反硝化作用便很难发生。但是在水生生态系统缺氧的时期,分子氮就可以通过反硝化过程而产生。科学家曾从某湖湖底和冰层下l米深处采集水样(冬季),注入标记氮的硝酸盐,然后将水样培养在与湖温相同的温度下。用质谱分析仪进行分析表明:湖底水样中的反硝化过程约比湖面附近快6倍,反硝化作用最重要的终结产物是分子氮,分子氮如果未在固氮活动中被重新利用则会返回大气圈库。
(5)氮的全球平衡
据估计,全球每年的固氮量为92×106吨(其中生物固氮54,工业因氮30,光化学固氮7.6和火山活动固氮0.2)。但是,借助于反硝化作用,全球的产氮量只有83×106吨(其中陆地43,海洋40和沉积层0.2)。两个过程的差额为9x106吨,这种不平衡主要是由工业固氮量的日益增长所引起的,所固定的这些氮是造成水生生态系统污染的主要因素。最近对海洋环境的研究表明,硝化作用大约可使海洋氮库补充20×106吨氮。从各种来源输入海洋的氮,大体上能被反硝化作用所平衡,基本土能维持一种稳定状态。
至今有一点是很清楚的,即氮的移动绝不是单方向、不可调节和与能量无关的。氮有很多循环路线,而每一条路线都受生物或非生物机制所调节,而且每一个过程都伴随着能量的消耗或释放。氮循环的这许多自我调节机制、反馈机制和对能量的依赖性曾导致提出了这样一个假设,即全球的氮循环是平衡的,固氮过程将被反硝化过程所抵消。目前这一假说还处在讨论之中。如果工业固氮量速率加速增长,而反硝化作用的增加速度又跟不上的话,那么任何已经达到的平衡都可能受到越来越大的压力。另外一个干扰因素是来自汽车和其他机动车所排放的二氧化氮,排放到大气中的含氮气体是造成空气污染的主要原因之一,而且这种污染物对于呼吸系统和大气臭氧层非常有害。
在某些生态系统内,硝化过程和反硝化过程能达到极好的协调,以有利于生态系统生产力的需要。这种协调过程在温带地区的冬季进行得最快,以致总是能保证在春季和夏初时使硝酸盐的数量达到最大,而这时也正是植物生长和繁殖需要硝酸盐最多的时候。
(引自《普通生态学》 高教社)
扩展资料
近20年我国森林固碳能力在渐强
《科学》杂志最近刊载的一篇文章在美国学术界引起强烈反响。北京大学北京大学城市与环境系教授方精云领导的研究小组,利用大量的野外实测资料,以及1949年至1998年的森林资源调查资料等,运用一种前所未有的方法,计算出中国森林近50年含碳量的变化,结果发现,前30年中国森林含碳量增加,后20年含碳量减少。这就是说,中国森林原来是像工厂的烟囱一样向大气中排放二氧化碳的,而现在正在将大气中的二氧化碳吸收回来,变成树木。并且吸收了多少碳,能够拿出数据。
人类活动每年释放大量二氧化碳,其中近一半增加了大气二氧化碳浓度,一部分被海洋吸收,而约有不到1/3的二氧化碳却“去向不明”。作为全球变化的前沿课题,近年来,这一著名的“二氧化碳去向不明”之谜强烈地吸引着各国科学家进行研究。
就像银行存储现金一样,森林可通过光合作用“存储”碳,而森林砍伐则释放碳。此消彼长间,我国森林究竟是吸收碳的“银行”,还是释放二氧化碳的“源泉”呢?
方精云教授等人历经近10年,研究了我国森林植被50年来碳存储量的动态变化。研究发现,20世纪70年代中期以前,由于毁林开荒等因素,我国森林向大气净排放了大量的二氧化碳。但在最近20年里情况发生逆转,我国森林净吸收二氧化碳的功能明显增强:每年工业排放出的二氧化碳平均有5%至8%也就是2600万吨被吸收。近20年共净吸收约4.5亿吨碳,相当于20世纪90年代中期中国工业二氧化碳年均排放量的一半。为缓解全球温室效应做出了积极贡献。在被“固定”的碳中,人工林占了80%。
“我国70年代末期以来倡导的植树造林工程,初衷本是防止水土流失和干旱,却出人意外地延缓了大气二氧化碳浓度的上升速度。”方精云说。据悉,我国人工林累计面积目前已居世界第一位,森林覆盖率也上升到16.55%。
国际同行高度评价我国科学家的研究结果。美国的《科学》杂志以及《国家地理杂志》都对这一研究做出了高度评价。《科学》杂志评论认为,中国科学家的工作为敏感的国家间及国际的森林及碳循环政策提供了一个基本原理。《科学》编委会首席指导萨格登说“它作为一个重要的里程碑,将会被长期引用,”
这一领域的资深专家、美国哈佛大学地球与行星科学系沃夫西教授说:“森林并不能奇迹般地阻止大气二氧化碳浓度的升高,但可在未来的几十年显著地缓和上升的速度。”哈佛大学的史蒂文教授认为“中国是一个最好的例子。如果中国能够做到,其他国家也应该做到。”
方精云教授认为:《科学》杂志刊登我们的论文正是看到了计算森林吸收碳还是释放碳的重要性。具体方法三言两语讲清楚不太容易。简单地说,就是根据每年木材生产量等统计数据和野外实测资料计算森林(包括根、茎和叶)有机碳的含量。如果这个含量增加了,就说明森林汇集了二氧化碳,如果减少了,就说明森林释放了二氧化碳。
生态学的传统观点认为,陆地生物圈对碳的吸收和排放是平衡的。因为动物的含碳量占整个陆地生物圈含碳量的比例很小,而森林占到90%以上,因此这一观点也就是说森林对碳的吸收和排放是平衡的。
但是在70年代末,美国科学家发现森林在释放碳,对于生态环境来说这可了不得,所以当时这是一条爆炸性的消息,于是美国生态学的精英都转而研究这一问题。1990年,美国国家航空航天局的一位专家发表森林正在吸收碳的观点。但持两种说法的专家都没有直接的证据。
现在我们的研究能够拿出具体的数据来说明森林在释放还是吸收碳的问题,这样就对两种争论谁对谁错做出判定了。
中国森林从释放碳到吸收碳的50年过程中有一个转折点,造成的主要原因是全球气温上升。如果砍伐树林,腐败的根、茎、叶等会向空气中释放二氧化碳,反之植树造林可以扭转这一趋势。
一个地区能够积累的碳的量多,就说明这个地区能够支持的社会经济能力就强。没有碳就没有生命,光合作用引起的碳的流动,会带动整个区域的生态系统正常运转起来。如果某个区(例如荒漠)碳的流动没有运转好,整个区的系统就无法正常运转。
所以我们绘制出了50年来我国960万平方公里土地上各个省的生产碳的能力图。这样可以知道什么地方生态系统支持社会经济的能力强,什么地方能力弱。
专家认为,我国科学家的发现,不仅为解释大气二氧化碳累积速度较低之谜提供了直接证据:工业排放的二氧化碳部分被森林所吸收,从而减轻了大气的压力;而且为环境外交事务提供了有力的技术支撑:为了减少全球二氧化碳的排放量,各国首脑签订了《京都议定书》,对发达国家排放二氧化碳的量做出了限制。而美国不久前退出《京都议定书》,其中的理由之一,就是认为中国、印度这样二氧化碳排放量大的发展中国家也应承担减排义务。但对于发展中国家来说,削减二氧化碳的排放意味着工厂的关闭,经济增长速度的减慢,失业率的增加。现在我们能够拿出科学的数据来说明森林吸收二氧化碳的量--占我国工业排放量的5%,对于经济发展来说,这是一个不小的数字。这个数字可以用于在削减二氧化碳的谈判上为我国的经济发展争取了更大的空间。
不过专家们强调,面对全球变暖,决不能只靠森林。正如帕卡拉所说,目前面临的最严峻挑战是,“必须控制化石燃料的排放量”。
扩展资料
森林与温室效应的新发现
温室效应被列为21世纪人类面临的最大威胁之一,它除了让天气变得越来越热外,还给全球的湿地沼泽、沿海低地、珊瑚礁、温带寒带大量物种带来毁灭性打击。
当人们恼于温室效应,地球变暖,而寄予厚望于森林吸收哦二氧化碳解救人类时,美国一群科学家的研究成果又给了人们劈头一棒。他们的最新研究表明:要想阻止气温逐年升高的趋势、化解温室效应的威胁,人类不能像从前那样寄厚望于森林,还得另想办法。因为森林吸收的大量二氧化碳,最终几乎会如数奉还给地球。
追根溯源,温室效应是过量的二氧化碳在大气中聚集而产生的一种天气现象。绿色植物进行光合作用吸收大量二氧化碳,这已是人们长期以来的共识,因而一直以来,森林被认为是解决温室效应的利器。这一最新研究表明,在大气二氧化碳浓度升高的情况下,森林吸收二氧化碳的能力可能不像人们想像的那么高,仅靠种树并不能有效地遏制全球变暖的趋势。改善我们的生存环境,减小温室效应,人类还要从减少二氧化碳的排放量做起。
5月24日出版的《自然》杂志发表了美国著名杜克大学的几位植物学家和生态学家的一份最新实验报告。7年前,这几位科学家在杜克大学校区附近选择了两片森林,开始了评估树木吸收二氧化碳能力的实验。据推算,到2050年,排放到大气中的二氧化碳量将比现在增加35%-50%。于是,他们就在第一组森林中不断施放二氧化碳浓度为560ppm(ppm为百万分之一)的气体,以模拟50年后的浓度水平;第二组森林则保持目前正常的二氧化碳水平,即浓度在365ppm左右。
在实验开始的前两年里,第一组森林的树木在高浓度二氧化碳下生长明显加快,生长速度比第二组森林的树木大约快25%。但两年后,生长速度却在很短的时间内迅速下降,最终和第二组森林的树木的生长速度基本持平。分析原因,主要是土壤中的养分,特别是氮养分随着树木的迅速生长而消耗殆尽。
实验结果显示,树木生长需要阳光、水分、养分等多种因素,缺一不可,森林可以在短期内加速吸收二氧化碳,但因为土壤中的养分无法配套供应,所以从长期来看,无法指望森林会消化掉50年后增加的50%的二氧化碳。另外,科学家们在跟踪观察二氧化碳在树木生长过程中的作用后发现,树木吸收的二氧化碳主要是对树叶的生长起作用,对树干的作用则不明显。这样带来的最要命的后果是,树叶脱落以后会慢慢腐烂,它所吸收的二氧化碳在3年内将几乎全部回到大气中,而并非像以前人们所说的那样,会留在土壤里。
主持这项实验的杜克大学生物学教授戴维・埃斯沃斯对这一结果评价说:“如果这一结果在全球植被生态中被普遍证实,人们就不得不对环境问题进行再思考了。”目前,已经有实验表明,热带树木与其他地区的树木相比,对二氧化碳的吸收能力相对更低一些。
外界对这份报告给予了极大关注。美国伊利诺伊大学的一位植物学家说,这一结果的“潜在重要性”会越来越明显,它彻底推翻了人们的传统观念。
尽管森林在涵养水源、防止水土流失、调节气候等方面都有无可替代的作用,但在大量消化二氧化碳的能力上,人们不能太乐观。参与这项实验的另一位生态学教授拉姆・奥林认为,这一实验结果的重要性就在于此。他说,也许人们只能从减少二氧化碳气体的排放量入手来解决温室效应问题了,这将使全球的环保问题变得更为复杂,利益冲突更为尖锐。
缅因州鲍登大学的科学家对火炬松的研究则发现,在二氧化碳浓度较高的环境里,落叶分解并释放出二氧化碳的速度也更快。树木吸收的碳有一半储藏在树叶中,树叶掉落后有的迅速分解、重新向大气中释放出二氧化碳,有的缓慢被土壤吸收。树叶分解速度加快,将使森林本身的碳沉降能力大打折扣。
在营养或温度等因素的影响下,环境变化甚至可能使一个生态圈由二氧化碳吸收者变成污染源。认为只要多种植树木就可以肆无忌惮地排放二氧化碳的想法,会导致决策者制定错误的环境政策,损害人类为遏制全球变暖作出的努力。
减少二氧化碳的排放量,一直是国际间一个极为敏感的问题。去年的海牙国际环保会议不欢而散,美欧之间争执的关键问题就是二氧化碳排放标准。美国坚持认为,对发达国家排放量的限制应当考虑到森林对二氧化碳的吸收因素。如果考虑这一因素,对50年后的前景就不必过于悲观,对发达国家的二氧化碳排放标准的要求就不必过于苛刻。现在,这项最新的实验成果可以说正击中了美国的要害。果真如此,美国将不能以森林为借口来逃避自己的责任,而全球只能进一步严格限制二氧化碳的排放量,这又将直接影响到世界各国经济的发展。
扩展资料
生态系统的基本功能(二)
三、生态系统的物质循环
生态系统中生命成分的生存和繁衍,除需要能量外,还必须从环境中得到生命活动所需要的各种营养物质。没有外界物质的输入,生命就停止,生态系统也将随之解体。物质还是能量的载体,没有物质,能量就会自由散失,也就不可能沿着食物链传递。所以,物质既是维持生命活动的结构基础,也是贮存化学能的运载工具。生态系统的能量流和物质流紧密联系,共同进行,维持着生态系统的生长发育和进化演替。两者的关系见图5-9。
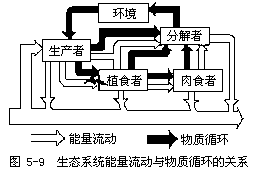
(一)物质循环的基本概念
生态系统中的物质主要指维持生命活动正常进行所必需的各种营养元素。这些物质也是通过食物链各营养级传递和转化的,从而构成了生态系统的物质流动。但物质循环却不是单方向性的。同一种物质可以在食物链的同一营养级内被生物多次利用。生态系统中各种有机物质经过分解者分解成可被生产者利用的形式归还环境中重复利用,周而复始地循环,这个过程叫做物质循环(Matteral cycle)。
生态系统物质循环研究常用到以下几个概念:
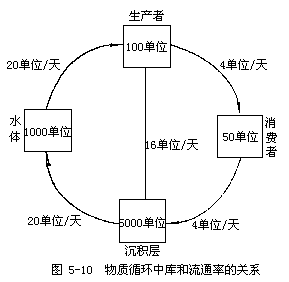
1.库(pools):是指某一物质在生物或非生物环境暂时滞留(被固定或贮存)的数量。例如,在一个湖泊生态系统中,磷在水体中的数量是一个库;磷在浮游植物中的含量又是一个库。磷在这两个库之间的动态变化就是磷这一营养物质的流动。可见,生态系统中的物质循环实际上就是物质在库与库之间的转移。根据库的大小和活跃程度常将其分为两类:(1)贮存库,其容积大而活动缓慢,一般为非生物成分,如岩石或沉积物;(2)交换库或循环库,营养物质在生物和其环境之间进行迅速交换的较小而又非常活跃的部分。如植物库,动物库,土壤库等。
2.流通率(Rate of circulation):是指单位时间、单位面积(或体积)内物质移动的量。可用克或公斤每亩・天来表示。
3.周转率(Turnover rates):是指某物质出入一个库的流通率与库量之比。即:
![]()
4.周转时间(Turnover time):周转率的倒数,即:
![]()
图5-10是一个池塘生态系统营养物质循环图示,可帮助理解上述概念及相互关系。
物质循环可在三个不同层次上进行:(1)生物个体,在这个层次上生物个体吸取营养物质建造自身,经过代谢活动又把物质排出体外,经过分解者的作用归还于环境;(2)生态系统层次,在初级生产者的代谢基础上,通过各级消费者和分解者把营养物质归还环境之中,故称为生物小循环或营养物质循环;(3)生物圈层次,物质在整个生物圈各圈层之间的循环,称生物地球化学循环。本节主要介绍后两种层次上的物质循环。生物小循环是在一个具体范围内进行的,其特点是物质流速快,周期短。生物地球化学循环简称生物地质大循环,其范围涉及到整个生物圈,并具有范围大,周期长和影响面广等特点。两种循环研究的侧重点有所不同。生物小循环侧重研究生态系统中营养物质的输入输出及其在各营养级间的交换过程。生物地质大循环则主要研究与人类生存密切相关的各种元素的全球性循环。两种循环是相互联系的,生物小循环不是封闭的,它受生物大循环的制约,是在生物地化大循环的基础上进行的。
(二)生态系统中营养物质的循环
在自然界已知的100多种化学元素中,生物正常生命活动所必需的约30-40种。这些元素在生物体的作用通常是不能相互替代的。生物对各种元素的需求量并不相同且有种间差异。据分析,细胞含有24种元素,其中C、H、O、N、P、S六种元素对生命起着特别重要的作用,大部分有机物分子是由这六种元素组成的。另外还有需求量较多的Ca、K、Na、Cl、Mg、Fe等六种元素,其它的元素是Mn、I、Mo、Co、Zn、Se、Cu、Cr、Sn、U、Si、F,这些元素含量很少,称为微量元素,但它们也是生命所不可缺少的。
生物所需要的这些营养物质的循环是在生态系统的四个基本成分之间进行的。另外,生态系统还可从降雨、空气流通和动物的迁入等不同途径使营养物质得到补充和更新。生态系统营养物质的整个循环过程中,生产者、分解者、水分和大气起着尤为重要的作用。生产者使无机物转变为有机物,分解者则把复杂有机物分解为生产者可重新利用的简单无机物。水和空气起介质作用,固体物只有溶于水中才能被生产者吸收利用。一些气态物和水分则需借助空气而由气孔等处进入生物体。
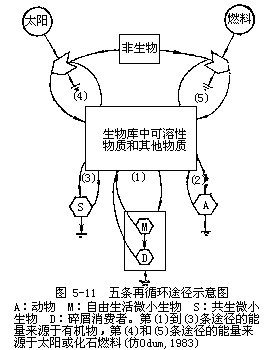
生态系统中营养物质再循环主要有以下几条途径:
1.物质由动物排泄返回环境。包括海洋等以浮游生物为优势种的水域生态系统都可能以这种途径为主。据研究(Harris,1959),浮游动物在其生存期间所排出的无机物和有机可溶性营养物质的数量,比它们死亡后经微生物分解所放出的数量要多好几倍,而且排泄的可溶性营养物能直接被生产者所利用。
2.物质由微生物分解碎屑过程而返回环境。在草原、温带森林及其它具有以碎屑食物链为主的生态系统,这种途径是主要的。
3.通过在植物根系中共生的真菌,直接从植物残体(枯枝落叶)中吸收营养物质而重新返回到植物体。在热带,尤其是热带雨林生态系统中存在着这种途径。
4.风化和侵蚀过程伴同水循环携带着沉积元素,由非生物库进入生物库。这是营养物质再循环的第四条途径。
5.动、植物尸体或粪便不经任何微生物的分解作用也能释放营养物质。如水中浮游生物的自溶可视为营养物质在生态系统中再循环的第五条途径。
6.人类利用化石燃料生产化肥,用海水制造淡水以及对金属的利用,可以认为是物质再循环第六条途径。
物质再循环的六条途径中,前五条是在自然状态下进行的。第六条途径的作用在加强,对生物圈的正常功能的影响也越来越大,由此引发许多问题正是环境生态学所研究的重要内容。生态系统中营养物质再循环的各种途径及其相互关系见图5-11。
(三)生物地球化学循环
生物地球化学循环是营养物质在生态系统之间的输入和输出,以及它们在大气圈、水圈和土壤圈之间的交换。根据物质参与循环时的形式,可将循环分为气相循环,液相循环和固相循环三种形式。
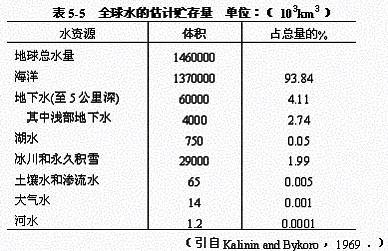
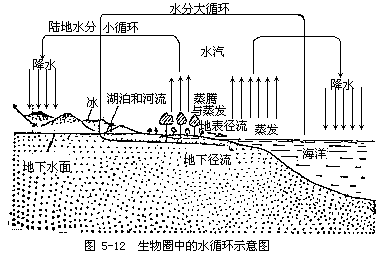
1.水循环:属液相循环,是在太阳能驱动下,水从一种形式转变为另一种形式,并在气流(风)和海流的推动下在生物圈内的循环。水在生物圈中的形式分为气态、液态和固态。各种形式水的数量及在地球上的分布见表5-5。
海洋和陆地上分布的水,由于太阳辐射作用,一部分蒸发为大气中的水汽,但陆地和海洋的蒸发量不相等。假如地球上的总降水量为100单位,那么,来源于海洋的蒸发量占84单位,来源陆地的只有16单位。但100单位的降水量中,海洋得77单位,陆地为23单位,海洋亏缺部分是由陆地入海的水而得到补偿。生物圈中水的循环过程见图5-12。
2.气相循环:亦称气体型循环(Gaseous cycles),循环物质为气态。以这种形态进行循环的主要营养物质有碳、氧、氮等。现以碳循环为例,说明这类循环的基本过程。
地球上碳的总量约为26×1015吨,绝大多数以无机形态存在于岩石圈中,大气中CO2约含碳7000亿吨,生物圈中碳的循环主要有三条途径:一是始于绿色植物并经陆生生物与大气之间的碳交换;第二条途径是海洋生物与大气间的碳交换;人类对化石燃料的应用是碳循环的第三条途径。生物圈中碳循环过程可简括为图5-13所示。
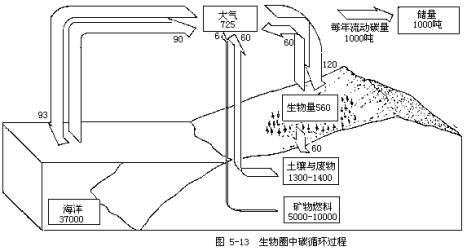
陆地和大气圈之间碳的交换几乎是平衡的,但人类的生产活动却不断地破坏着这种平衡。据估计,人为因素可使大气中的CO2每年增加7.5×109吨,这仅是人类向大气释放CO2数量的1/3,其余 2/3则被海洋和陆地植物所吸收。CO2能阻止地球热量的散失,导致地球气温的升高,成为当今全世界所忧虑的重要环境问题之一。
3.固相循环或称沉积型循环:参与循环的物质中很大一部分又通过沉积作用进入地壳而暂时或长期离开循环。这是一种不完全的循环,属于这种循环方式的有磷、钾和硫等,现以磷循环为例介绍其循环过程。
磷主要贮存于岩石和天然磷酸矿中,它的循环与水循环密切相关。磷必须形成可溶性磷酸盐才能进入循环。首先,存在于岩石和天然磷酸盐矿床中的磷,通过风化淋溶、侵蚀等作用及采矿而被释放出来,进入水圈和土壤圈。溶于水后形成可溶性磷酸盐。植物从环境中吸收磷,然后沿食物链在生态系统中传递并经分解者作用又归还到环境中,可被植物再次利用。在陆地生态系统中,土壤中的磷易于与钙、铁结合而形成不能为植物吸收利用的难溶性钙盐。陆地上的一部分磷可随水进入江河,湖泊和海洋,大部分以磷酸钙的形式沉积于海底或珊瑚岩中。水域生态系统中,从浮游植物吸收无机磷开始,浮游植物被浮游动物或食碎屑生物所食,与陆地一样沿食物链传递。但浮游动物代谢排出的无机态磷可被浮游植物再次利用。所以水体中磷的周转速度比陆地快。水体中的有机磷可被微生物所分解和利用,并能再次转变为无机态而被植物利用。但水体中也有相当一部分磷在浅水或深水中沉积下来(图5-14)。

磷循环的这种不完全性,使土壤中的含量因农作物的吸收利用而不断减少,常成为作物生长发育的限制因素。
四、生态系统中的信息传递
生态系统的功能整体性除体现在生物生产过程,能量流动和物质循环等方面外,还表现在系统中各生命成分之间存在着信息传递,习惯上称为信息流(Information flow)。这是生态系统生态学研究中的一个薄弱环节,同时也是一个颇具吸引力的研究领域,尤其是近几年关于行为信息的研究进展更为迅速。
从信息传递的角度看,生态系统中的各种信息主要可分为两大类:
(一)物理信息:以物理过程为传递形式的信息,如光、声音、颜色等。动物的求偶行为、恐吓、报警行为等都与物理信息有关。例如,鸟类在繁殖季节时,常伴有鲜艳色彩的羽毛或其它的奇特装饰以及美妙动人的鸣叫等,各种“特长”都在求偶时尽情显露。雄孔雀开屏的表演能促使雌孔雀自动地接近它,并摆出接受爱抚的某种姿态,而雄孔雀的表演动作也就能迅速地促使两者结为伴侣。
(二)化学信息:生物代谢产生的一些物质,尤其是各种腺体分泌的各类激素等均属传递信息的化学物质。同种动物间以释放化学物质传递信息是相当普遍的现象。如长爪沙鼠能从四种小型啮齿类的外激素中分出同种的气味;属(clethrionomys)可通过气味区别出本亚种和其它亚种。有些动物没有固定的领域,但它们却可利用特定方式交换情报,以调整区域的合理利用。如猎豹和猫科动物有着高度特化的用尿标志的结构,它们总是仔细观察前兽留下来的痕迹,并由此传达时间信息,避免与栖居同一地区的对手相互遭遇。生物种间也存在着化学通讯联系,而且这种联系不仅见于动物与动物之间,也常见于动物与植物间,植物与植物之间。物种在进化过程中,逐渐形成释放化学信号于体外的特性。这些信号或对释放者本身有利,或有益于信号接受者。它们影响着生物的生长、健康或物种生物特征。黄鼬(黄鼠狼)有一种嗅腺,释放出来的臭液气味难闻,它既有防止敌害追捕的作用,也有助于获取食物。再如,烟草中的尼古丁和其它植物碱可使烟草上的蚜虫麻痹;成熟橡树叶子含有的单宁不仅能抑制细菌和病毒,同时它还使蛋白质形成不能消化的复杂物质,限制脊椎动物和蛾类幼虫的取食;胡桃树(Jugla ns SP.)的叶表面可产生一种物质,被雨水冲洗落到土壤中,可抑制土壤中其它灌木和草本植物的生长。这些都是植物自我保护向其它生物所发出的化学信息。
尽管生态系统信息流的研究还存在许多困难,但生物间的这种通讯联系的作用对生态系统的影响是十分明显的,特别是化学信息物质的作用更为重要。在一个生态系统中,化学信息物质的破坏常导致群落成分的变化。同时它们还影响着群落的营养及空间结构和生物间的彼此联系。因为各种信息的作用不是孤立的,而是相互制约、互为因果关系。另外,通过对生物信息传递的研究,还可以获得其它的生态信息。例如,狼也是用尿标记活动路线的动物,它们常用树桩、树木等作为“气味站”,在开阔地带,任何一突起物都可以被狼选择为标记对象。有时一群狼依次排尿于同一标记处。在冬季,这种标记站常形成相当大的冰坨。人们可通过对冰坨的分析获得狼群大小和数量的信息。
在生物中,用作通讯的形式和与之相连的感觉器官是多种的,据此可将生态系统中的信息流分成三类:1)化学通讯信息流;即由嗅觉和味觉通路传导信息;2)机械通讯信息流:经触觉、听觉通路进行信息传导;3)辐射通讯信息流:由光感受或视觉来执行其通讯机能。甚至超声波和电场也被生物用来传递信息。
金岗,环境生态学,高等教育出版社
扩展资料
生态系统与生态循环
(1) 生态系统的概念
生态系统是生态学的一个概念。生态学是一门研究生物和其生活环境的相互关系的科学,是生物学的主要分支。
一个物种在一定空间范围内的所有个体的总和在生态学里称为种群(population),所有不同种的生物的总和为群落(community),生物群落连同其所在的物理环境共同构成生态系统(ecosystem)。生态系统就是生命系统和环境系统在特定空间的组合,其特征是系统内部以及系统与系统外部之间存在着能量的流动和由此推动的物质的循环。例如,森林、草原、河流、湖泊、山脉或其一部分都是生态系统;农田、水库、城市则是人工生态系统。生态系统具有等级结构,即较小的生态系统组成较大的生态系统,简单的生态系统组成复杂的生态系统,最大的生态系统是生物圈。
(2) 生态系统的结构
任何一个生态系统都由生物群落和物理环境两大部分组成。阳光、氧气、二氧化碳、水、植物营养素(无机盐)是物理环境的最主要要素,生物残体(如落叶、秸杆、动物和微生物尸体)及其分解产生的有机质也物理环境的重要要素。物理环境除了给活的生物提供能量和养分之外,还为生物提供其生命活动需要的媒质,如水、空气和土壤。而活的生物群落是构成生态系统精密有序结构和使其充满活力的关键因素,各种生物在生态系统的生命舞台上各有角色。
生态系统的生命角色有三种,即生产者、消费者和分解者,分别由不同种类的生物充当。生产者吸收太阳能并利用无机营养元素(C、H、O、N等)合成有机物,将吸收的一部分太阳能以化学能的形式储存在有机物中。生产者的主体是绿色植物,以及一些能够进行光合作用的菌类。由于这些生物能够直接吸收太阳能和利用无机营养成分合成构成自身有机体的各种有机物,我们称它们是自养生物。消费者是直接或间接地利用生产者所制造的有机物作为食物和能源,而不能直接利用太阳能和无机态的营养元素的生物,并最终还原为植物可以利用的营养物。消费者和分解者都不能够直接利用太阳能和物理环境中的无机营养元素,我们称它们为异养生物。值得特别指出的是,物理环境(太阳能、水、空气、无机营养元素)、生产者和分解者是生态系统缺一不可的组成部分,而消费者是可有可无的。这一点可以在图2-2 中得到直观的反映。
(3) 生态系统的物质循环
在生态系统中,物质从物理环境开始,经生产者、消费者和分解者,又回到物理环境,完成一个由简单无机物到各种高能有机化合物,最终又还原为简单无机物的生态循环。通过该循环,生物得以生存和繁衍,物理环境得到更新并变得越来越适合生物生存的需要。在这个物质的生态循环过程中,太阳能以化学能的形式被固定在有机物中,供食物链上的各级生物利用。
生物维持生命所必需的化学元素虽然为数众多,但有机体的97%以上是由氧、碳、氢、氮和磷五种元素组成的。作为物质循环的例子,下面分别介绍碳、氮和磷的生态循环过程。
① 碳循环
碳是构成生物原生质的基本元素,虽然它在自然界中的蕴藏量极为丰富,但绿色植物能够直接利用的仅仅限于空气中的二氧化碳(CO2)。生物圈中的碳循环主要表现在绿色植物从空气中吸收二氧化碳,经光合作用转化为葡萄糖,并放出氧气(O2)。在这个过程中少不了水的参与。有机体再利用葡萄糖合成其他有机化合物。碳水化合物经食物链传递,又成为动物和细菌等其他生物体的一部分。生物体内的碳水化合物一部分作为有机体代谢的能源经呼吸作用被氧化为二氧化碳和水,并释放出其中储存的能量。由于这个碳循环,大气中的CO2大约20年就完全更新一次。
② 氮循环
在自然界,氮元素以分子态(氮气)、无机结合氮和有机结合氮三种形式存在。大气中含有大量的分子态氮。但是绝大多数生物都不能够利用分子态的氮,只有象豆科植物的根瘤菌一类的细菌和某些蓝绿藻能够将大气中的氮气转变为硝态氮(硝酸盐)加以利用。植物只能从土壤中吸收无机态的铵态氮(铵盐)和硝态氮(硝酸盐),用来合成氨基酸,再进一步合成各种蛋白质。动物则只能直接或间接利用植物合成的有机氮(蛋白质),经分解为氨基酸后再合成自身的蛋白质。在动物的代谢过程中,一部分蛋白质被分解为氨、尿酸和尿素等排出体外,最终进入土壤。动植物的残体中的有机氮则被微生物转化为无机氮(氨态氮和硝态氮),从而完成生态系统的氮循环。
③ 磷循环
磷是有机体不可缺少的元素。生物的细胞内发生的一切生物化学反应中的能量转移都是通过高能磷酸键在二磷酸腺苷(ADP)和三磷酸腺苷(ATP)之间的可逆转化实现的。磷还是构成核酸的重要元素。磷在生物圈中的循环过程不同于碳和氮,属于典型的沉积型循环。生态系统中的磷的来源是磷酸盐岩石和沉积物以及鸟粪层和动物化石。这些磷酸盐矿床经过天然侵蚀或人工开采,磷酸盐进入水体和土壤,供植物吸收利用,然后进入食物链。经短期循环后,这些磷的大部分随水流失到海洋的沉积层中。因此,在生物圈内,磷的大部分只是单向流动,形不成循环。磷酸盐资源也因而成为一种不能再生的资源。
(4) 生态系统的能量流
推动生物圈和各级生态系统物质循环的动力,是能量在食物链中的传递,即能量流。与物质的循环运动不同的是,能量流是单向的,它从植物吸收太阳能开始,通过食物链逐级传递,直至食物链的最后一环。在每一环的能量转移过程中都有一部分能量被有机体用来推动自身的生命活动(新陈代谢),随后变为热能耗散在物理环境中。
为了反映一个生态系统利用太阳能的情况,我们使用生态系统总产量这一概念。一个生态系统的总产量是指该系统内食物链各个环节在一年时间里合成的有机物质的总量。它可以用能量、生物量表示。生态系统中的生产者在一年里合成的有机物质的量称为该生态系统的初级总产量。在有利的物理环境条件下,绿色植物对太阳能的利用率一般在1%左右。生物圈的初级生产总量约4.24′1021焦耳/年,其中海洋生产者的总产量约1.83′1021焦耳/ 年,陆地的约为2.41′1021焦耳/年。总产量的一半以上被植物的呼吸作用所耗用,剩下的称为净初级产量。各级消费者之间的能量利用率也不高,平均约为10%,即每经过食物链的一个环节,能链的净转移率平均只有十分之一左右。因此,生态系统中各种生物量按照能量流的方向沿食物链递减,处在最基层的绿色植物的量最多,其次是草食动物,再次为各级肉食动物,处在顶级的生物的量最少,形成一个生态金字塔。只有当生态系统生产的能量与消耗的能量大致相等时,生态系统的结构才能维持相对稳定状态,否则生态系统的结构就会发生剧烈变化。